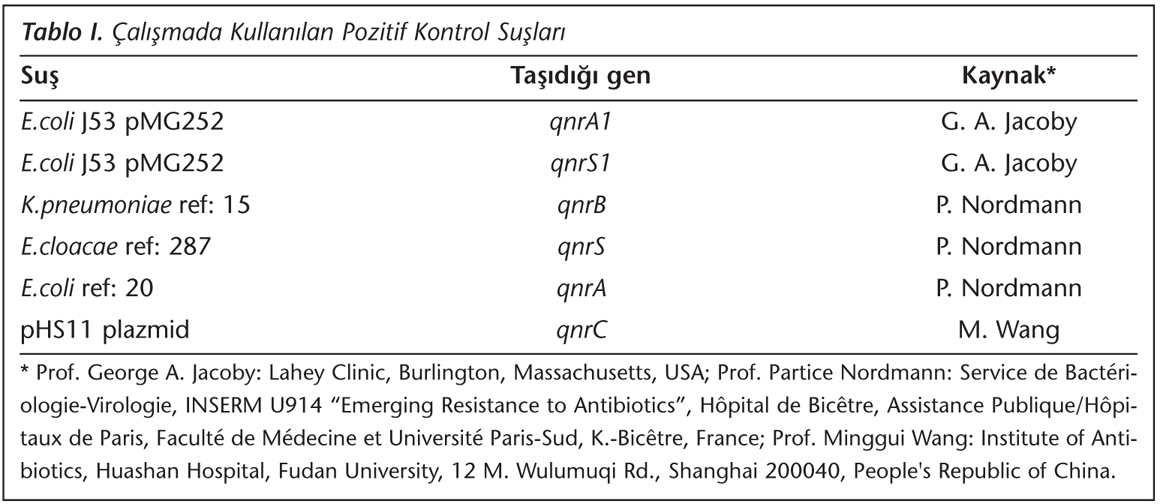
Enterobacteriaceae Üyelerinde Plazmid
Aracılı Kinolon Direnç Determinantlarının Araştırılması:
Çok Merkezli Bir Çalışma
Investigation
of Plasmid-Mediated Quinolone Resistance
Determinants in Enterobacteriaceae: A Multicenter Study
Ahmet
Yılmaz ÇOBAN1, Okan Kadir NOHUT2, Yeliz TANRIVERDİ ÇAYCI1, Gülçin BAYRAMOĞLU3,
Müjgan PİRİNÇÇİLER4, Ebru ÇETİNKAYA5, Çiğdem ÇEKİÇ CİHAN6, Bülent BOZDOĞAN7,
Belma DURUPINAR1
1 Ondokuz Mayıs Üniversitesi Tıp Fakültesi, Tıbbi Mikrobiyoloji Anabilim Dalı, Samsun.
1 Ondokuz Mayıs University Faculty of Medicine, Department of Medical Microbiology, Samsun, Turkey.
2 Ondokuz Mayıs Üniversitesi Fen Edebiyat Fakültesi, Biyoloji Bölümü, Samsun.
2 Ondokuz Mayıs University Faculty of Science and Literature, Department of Biology, Samsun, Turkey.
3 Karadeniz Teknik Üniversitesi Tıp Fakültesi, Tıbbi Mikrobiyoloji Anabilim Dalı, Trabzon.
3 Karadeniz Technical University Faculty of Medicine, Department of Medical Microbiology, Trabzon, Turkey.
4 Çanakkale Devlet Hastanesi, Klinik Mikrobiyoloji Laboratuvarı, Çanakkale.
4 Canakkale State Hospital, Clinical Microbiology Laboratory, Canakkale, Turkey.
5 Dr. Nafiz Kören Sincan Devlet Hastanesi, Klinik Mikrobiyoloji Laboratuvarı, Ankara.
5 Dr. Nafiz Koren Sincan State Hospital, Clinical Microbiology Laboratory, Ankara, Turkey.
6 Çorum Göğüs Hastalıkları Hastanesi, Klinik Mikrobiyoloji Laboratuvarı, Çorum.
6 Corum Chest Diseases Hospital, Clinical Microbiology Laboratory, Corum, Turkey.
7 Adnan Menderes Üniversitesi Tıp Fakültesi, Tıbbi Mikrobiyoloji Anabilim Dalı, Aydın.
7 Adnan Menderes University Faculty of Medicine, Department of Medical Microbiology, Aydin, Turkey.
ÖZET
1986 yılından beri kullanımda olan florokinolonlar hem gram-pozitif hem de gram-negatif bakterilere etkili ajanlardır. Kinolonlar, DNA giraz ve topoizomeraz IV enziminin inhibisyonu sonucu bakterisidal etki gösterir. Kinolonlara direnç gelişmesinde başlıca mekanizmalar bu enzimlerde meydana gelen kromozomal mutasyonlar ve atım pompaları veya dış zar geçirgenliğinde azalma nedeniyle hücre içinde ilaç birikiminin azalmasıdır. Ancak son yıllarda bu direnç mekanizmalarına ek olarak ortaya çıkan bir mekanizma da plazmid aracılı kinolon direncidir. Bu direnç genleri qnr olarak adlandırılmıştır. Qnr genleri tek başlarına kinolon direncine neden olmasa da kinolon duyarlılığında azalmaya ve minimum inhibitör konsantrasyonu değerlerinde artmaya neden olmaktadır. Günümüzde tanımlanmış beş qnr gen bölgesi (qnrA, qnrB, qnrC, qnrD ve qnrS) mevcuttur. Bu çalışmada plazmid aracılı kinolon direncini saptamak amacıyla, Türkiye'nin dört farklı bölgesinden toplanan Enterobacteriaceae ailesine ait klinik izolatlarda qnrA, qnrB, qnrC ve qnrS genlerinin varlığı araştırılmıştır. Çalışmaya, Mayıs 2009-Temmuz 2009 tarihleri arasında dört farklı merkezden [Trabzon (n= 387), Çanakkale (n= 82), Ankara (n= 96), Tokat (n= 82)] toplanmış olan Enterobacteriaceae ailesine ait 647 bakteriyel izolat dahil edilmiş ve bu suşlarda qnrA, qnrB, qnrC ve qnrS genleri multipleks polimeraz zincir reaksiyonu (PCR) yöntemiyle araştırılmıştır. Çalışmamızda, 2 izolatta qnrA, 12 izolatta qnrB, 4 izolatta qnrC ve 10 izolatta qnrS pozitifliği saptanmış ve pozitifliklerin doğrulaması amacıyla dizi analizi yapılmıştır. Dizi analizi sonucunda pozitif izolat sayısı azalmış; iki izolatta qnrA1 [Enterobacter cloacae (kod no. 796), Salmonella grup B (kod no. 491)], iki izolatta qnrB1 [Salmonella grup B (kod no. 491), Citrobacter freundii (kod no. 768)], bir izolatta qnrB6 [Escherichia coli (kod no. CC1800)], bir izolatta qnrB9 [E.coli (kod no. CC1873)], bir izolatta qnrB24 [Citrobacter koseri (kod no. MP5200)], bir izolatta qnrB27 [C.freundii (kod no. 842)], iki izolatta qnrS1 [E.coli (kod no. CC1705), E.coli (kod no.159)] ve bir izolatta qnrB2 [E.coli (kod no. 843)] pozitifliği saptanmıştır. Qnr geni saptanan izolatlardan biri siprofloksasine, ikisi nalidiksik aside dirençli bulunmuştur. Mutasyona bağlı kinolon direncinin klinik suşlar arasında artmasının yanı sıra, aktarılabilir kinolon direncinin yaygınlaşması klinik açısından önemli sonuçlar doğurabilecektir. Bu nedenle aktarılabilen kinolon direncinin izlenmesi ve yayılım oranlarının saptanması, alınacak önlemlerin belirlenmesi için önemlidir.
Anahtar sözcükler: Qnr; plazmid; Enterobacteriaceae; florokinolon; direnç; Türkiye.
ABSTRACT
Fluoroquinolones which are in use since 1986, are effective agents both against gram-positive and gram-negative bacteria. Quinolones show bactericidal effect as a result of inhibition of DNA gyrase and topoisomerase IV enzymes. Main quinolone resistance mechanisms are chromosomal mutations in these enzymes and decreased intracellular accumulation due to efflux pumps or decreased membrane uptake. Recently a new quinolone resistance mechanism mediated by plasmids has been defined. These plasmids carry genes called as qnr. Qnr genes do not cause quinolone resistance but they cause decreased quinolone susceptibility and lead to higher minimum inhibitory concentrations. Currently there are qnrA, qnrB, qnrC, qnrD and qnrS genes. This study was aimed to investigate the presence of plasmid-mediated quinolone resistance determinants in Enterobacteriaceae isolates collected from four different centers in Turkey. A total of 647 isolates (387 from Trabzon, Black Sea region; 82 from Canakkale, Trace region; 96 from Ankara, Central Anatolia region; 82 from Tokat, Black Sea region) belonging to the Enterobacteriaceae family collected between May-July 2009, were included in the study. Presence of qnrA, qnrB, qnrS and qnrC genes were investigated by multiplex polymerase chain reaction (PCR) method and confirmed by gene sequencing. The results of the PCR amplification revealed that 2 isolates were positive for qnrA, 12 isolates were positive for qnrB, 4 isolates were positive for qnrC and 10 isolates were positive for qnrS. However, the number of positive strains decreased with the use of gene sequencing, and this method led to the identification of qnrA1 in two isolates [Enterobacter cloacae (code. 796), Salmonella group B (code. 491)], qnrB1 in two isolates [Salmonella group B (code. 491), Citrobacter freundii (code. 768)], qnrB6 in one isolate [Escherichia coli (code. CC1800)], qnrB9 in one isolate [E.coli (code. CC1873)], qnrB24 in one isolate [Citrobacter koseri (code. MP5200)], qnrB27 in one isolate [C.freundii (code. 842)], qnrS1 in two isolates [E.coli (code. CC1705), E.coli (code.159)] and qnrB2 in one isolate [E.coli (code. 843)]. One of the isolates that carried the qnr gene was ciprofloxacin-resistant and two isolates were nalidixic-acid resistant. Transferable quinolone resistance due to the dissemination of qnr genes may have important impacts in terms of infection control and treatment problems. Survey of plasmid mediated quinolone resistance will help to determine the size of the issue and guide the measures that should be taken to avoid escalation of resistance and dissemination problem.
Key words: Qnr; plasmid; Enterobacteriaceae; fluoroquinolone; resistance; Turkey.
Geliş Tarihi (Received): 18.11.2011 • Kabul Ediliş Tarihi (Accepted): 23.02.2012
GİRİŞ
Florokinolonlar uzun yıllardır hem tıpta hem de veterinerlikte kullanılan antimikrobiyal ajanlardır1. 1962 yılında ilk keşfedilen kinolon olan nalidiksik asidin, sadece üriner sistem enfeksiyonu etkeni olan gram-negatif basiller üzerine etkili olması kullanımını sınırlandırmıştır2. Zaman içerisinde yapılan çalışmalarla altıncı atoma florun eklenmesiyle aktiviteleri artmış ve etki spektrumları genişlemiştir. Siprofloksasin özellikle Pseudomonas aeruginosa üzerine olan etkinliği nedeniyle gram-negatif bakteriyel enfeksiyonlarda, levofloksasin de Streptococcus pneumoniae ve atipik pnömoni etkenleri olan Mycoplasma pneumoniae, Chlamydia pneumoniae ve Legionella pneumophilia üzerine olan etkinliği nedeniyle solunum yolu enfeksiyonlarında sıklıkla kullanılmaktadır3,4. Yaygın kullanıma bağlı olarak kinolonlara karşı direnç gelişimi de hızla artmaktadır.
Kinolonlara karşı direnç gelişiminde en önemli mekanizmalardan biri DNA giraz (gyrA ve gyrB) ve topoizomeraz IV'te (parC ve parE) meydana gelen mutasyonlardır. Buna ilave olarak dışa atım pompalarıyla ilacın dışarı atımı ve dış membran geçirgenliğinde azalma da kinolonlara karşı önemli direnç gelişim mekanizmalarıdır4. Bu direnç mekanizmalarına, ilk kez 1998 yılında bir Klebsiella pneumoniae klinik izolatında tespit edilen yeni bir direnç mekanizması daha eklenmiş; bu direncin plazmidler aracılığıyla aktarıldığı saptanmış ve bu gen bölgesi qnr olarak adlandırılmıştır5. Ayrıca, yine plazmidle aktarılan kinolon atım pompası qepA, çoklu ilaç direnci atım pompası oqxAB ve modifiye aminoglikozid asetil transferaz geni aac(6')-Ib-cr tanımlanmıştır4,6,7. Plazmidle aktarılan qnr genlerinin ilk tespitinden bugüne kadar farklı alt tipler (yedi adet qnrA, 27 adet qnrB, dört adet qnrS, birer adet qnrC ve qnrD geni) bildirilmiştir7,8.
Qnr determinantları florokinolonlara karşı sadece duyarlılıkta azalmaya yol açmakta ve minimum inhibitör konsantrasyonu (MİK) değerlerinde 16-32 kat artışa neden olabilmektedir9. Plazmid aracılı kinolon direnci, Enterobacteriaceae ailesinde giderek artan sıklıkta dünyanın farklı bölgelerinden bildirilmektedir10. Bu çalışmada plazmid aracılı kinolon direncini saptamak amacıyla, Türkiye'nin dört farklı bölgesinden gelen Enterobacteriaceae ailesine ait klinik izolatlarda qnrA, qnrB, qnrC ve qnrS genlerinin varlığı araştırılmıştır.
GEREÇ ve YÖNTEM
Bakteriyel İzolatlar
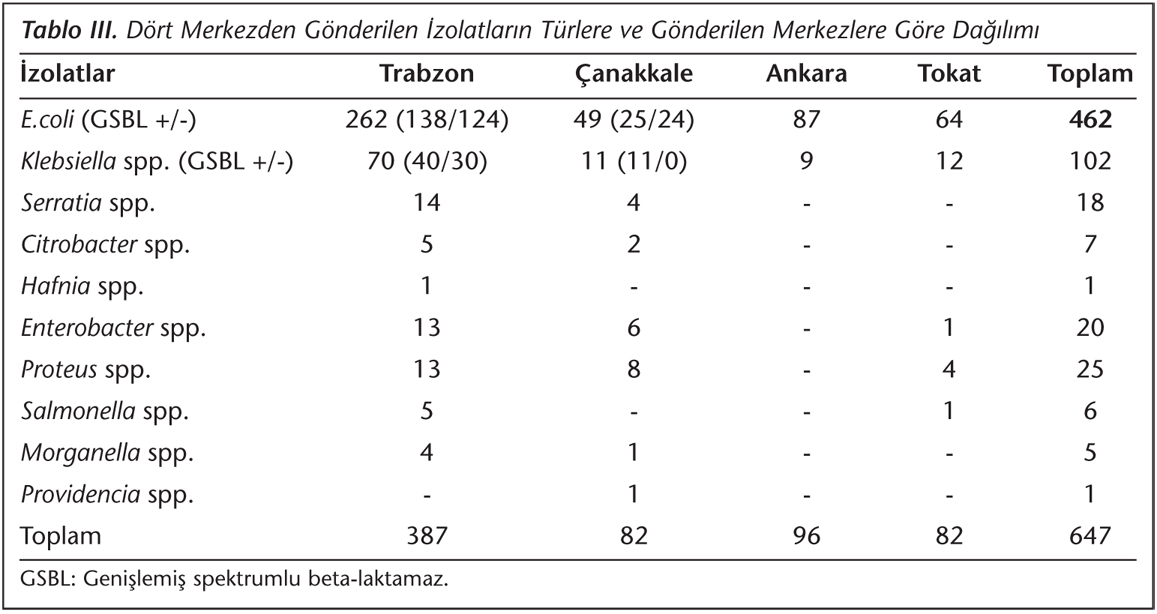
Çalışmaya, Mayıs 2009-Temmuz 2009 tarihleri arasında dört farklı merkezden (Trabzon, Çanakkale, Ankara, Tokat) toplanmış olan Enterobacteriaceae ailesine ait 647 bakteriyel izolat dahil edildi. Karadeniz Teknik Üniversitesi Tıp Fakültesi, Tıbbi Mikrobiyoloji Anabilim Dalından gelen 387 izolat Vitek2 Compact cihazı (bioMeriux, Fransa); Çanakkale Devlet Hastanesi Klinik Mikrobiyoloji Laboratuvarından gelen 82 izolat Trek Diagnostic Sensititre (Trek Diagnostic Systems, ABD) cihazı; Ankara Dr. Nafiz Kören Sincan Devlet Hastanesi Klinik Mikrobiyoloji Laboratuvarından gelen 96 izolat BBL Crystal ID (BD, ABD) cihazı; Tokat Doğumevi Hastanesinden gelen 82 izolat ise konvansiyonel yöntemler ile tanımlandı. İzolatların antibiyotik duyarlılıkları disk difüzyon yöntemiyle test edildi.
Polimeraz zincir reaksiyonu (PCR) ve sekans işlemi sonucunda qnr geni taşıdığı tespit edilen izolatların siprofloksasin ve nalidiksik asit MİK değerleri sıvı mikrodilüsyon yöntemiyle saptandı.
Polimeraz Zincir Reaksiyonu (PCR)
DNA ekstraksiyonu kaynatma yoluyla yapıldı. Bu işlemde; bir gün öncesinden Mueller-Hinton agar besiyerinde üretilen bakterilerden birkaç koloni alınıp, 500 µl steril distile su içinde süspanse edildi ve 100°C'de 15 dakika kaynatıldı. Daha sonra 15000xg'de 20 dakika santrifüj işleminden sonra süpernatan, PCR'de kalıp DNA olarak kullanılmak üzere alındı. PCR yönteminde kontrol olarak; qnrA, qnrB, qnrC ve qnrS genlerini taşıyan pozitif suşlar (Tablo I) ve ayrıca DNA içermeyen bir negatif kontrol kullanıldı.
PCR işleminde qnrA, qnrB, qnrC ve qnrS gen bölgeleri multipleks olarak çalışıldı. PCR'de Kim ve arkadaşları11 tarafından daha önce tanımlanan primer çiftleri kullanıldı (Tablo II). Amplifikasyon programı; 95°C'de 1 dakika; 35 siklustan oluşan 95°C'de 1 dakika, 60°C'de 1 dakika ve 72°C'de 1 dakika ve son uzama aşaması olarak 72°C'de 10 dakika olacak şekildeydi. PCR ürünlerine %2 agaroz jelde 120V, 60 dakika 1xTBE tamponda elektroforez işlemi uygulandı. Daha sonra 20 dakika 0.05 mg/L etidyum bromür ile boyanarak görüntülendi. PCR işlemi sonunda pozitif olduğu düşünülen izolatlara tekrar tek primer ile PCR uygulandı.

Tekrarlanan PCR sonrası pozitif gen bölgesi saptanan izolatlar dizi analizi ile doğrulandı. Bunun için amplikonlar Macrogen (Güney Kore) firmasına gönderildi. Dizi analizi sonuçları NCBI Nucleotide Blast programı kullanılarak Gen Bankasıyla karşılaştırıldı.
BULGULAR
Çalışmaya alınan bakteriyel izolatların tür ve merkezlere göre dağılımı Tablo III'de verilmiştir. E.coli izolatlarının %35.3 (163/462)'ünde, Klebsiella spp. izolatlarının ise %50 (51/102)'sinde genişlemiş spektrumlu beta-laktamaz (GSBL) pozitiftir. Qnr geni saptanan izolatlardan ise sadece qnrS1 saptanan 159 no'lu E.coli izolatında GSBL pozitif saptanmıştır. Qnr geni saptanan izolatların siprofloksasin ve nalidiksik asit MİK değerleri Tablo IV'te görülmektedir.
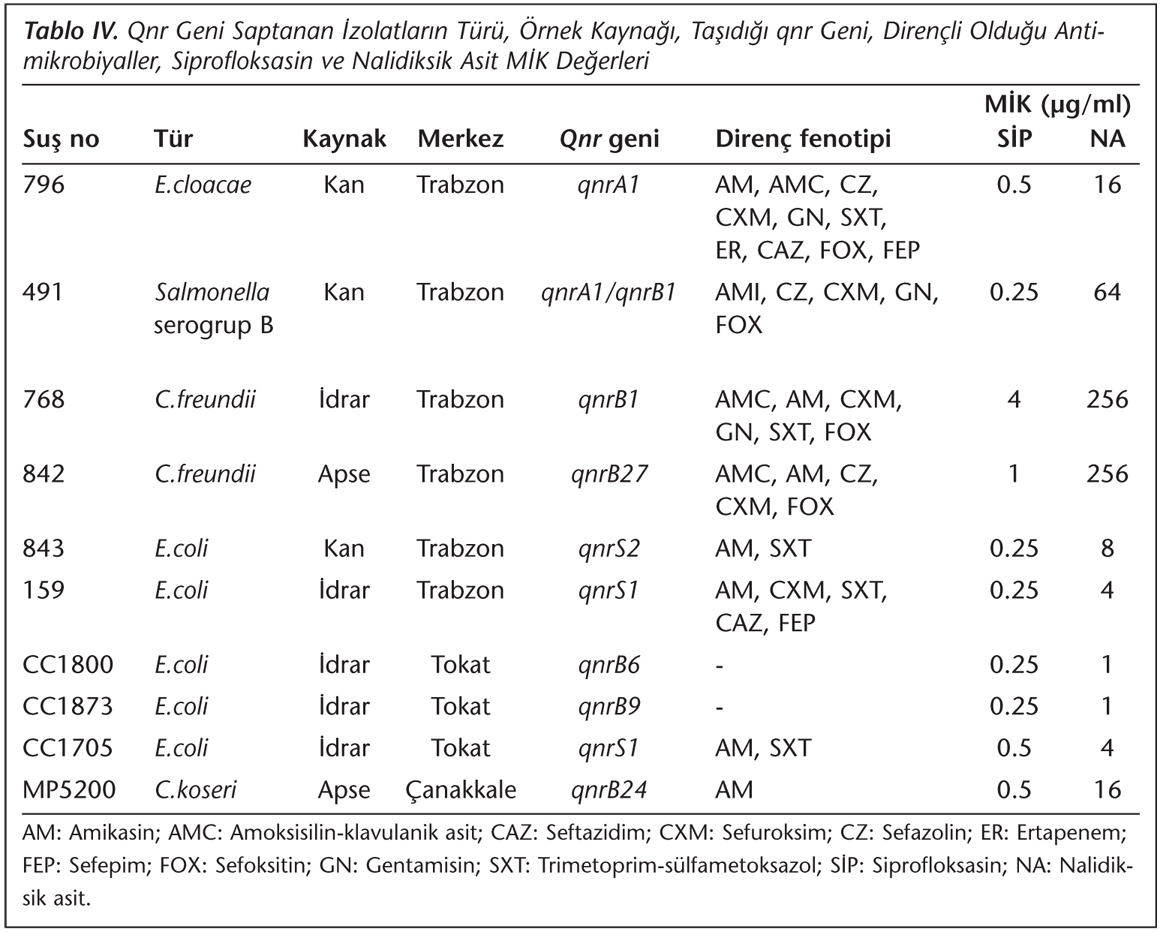
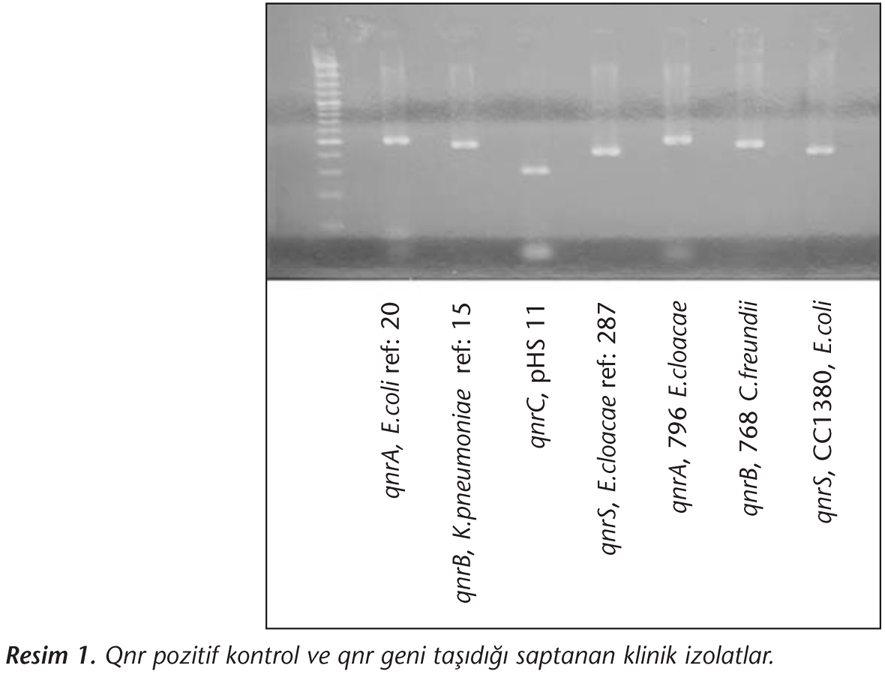
Multipleks PCR işlemi sonunda 99 izolatta qnrA, 157 izolatta qnrB, 17 izolatta qnrC ve 46 izolatta qnrS şüpheli pozitif olarak belirlenmiş; ancak sonrasında pozitif olan izolatlarda tekrarlanan PCR ile 2 izolatta qnrA, 12 izolatta qnrB, 4 izolatta qnrC ve 10 izolatta qnrS pozitifliği saptanmıştır. Doğrulama amacıyla yapılan dizi analizi sonucunda ise pozitif bulunan izolat sayısı 10'a düşmüş, bir izolatın (491 no) hem qnrA1 hem de qnrB1 geni taşıdığı izlenmiştir (Tablo IV). İzolatlarda tespit edilen qnr tipleri Tablo IV'te, qnr gen bölgesi saptanan izolatlar ve pozitif kontrol suşlarına ait görüntü ise Resim 1'de verilmiştir. Bu sonuçlara göre çalışmada saptanan qnr oranları sırasıyla; qnrA %0.3, qnrB %0.9, qnrS %0.4'tür.
TARTIŞMA
Kinolonlar bakterisidal etkiye sahip sentetik kemoterapötik ajanlardır. 1960'lı yıllarda kullanıma giren nalidiksik asitten günümüze kimyasal yapılarında yapılan değişikliklerle hem antimikrobiyal etki spektrumu genişlemiş hem de farmakodinamik özellikleri değişmiştir. Bugün hem gram-pozitif hem gram-negatif hem de anaeroplar üzerine etkilidirler. Kinolonların zaman içerisinde meydana gelen bu değişikliklerle birlikte kullanımları da yaygınlaşmıştır4. Yaygın kullanım direnç sorununu da birlikte getirmiştir. Kinolon grubu antibiyotiklere başlıca direnç gelişimi iki mekanizmayla olmaktadır; i) kinolonların hedefindeki değişim ve ii) membran geçirgenliğinde azalma veya atım pompalarının varlığına bağlı olarak hücre içinde ilaç birikiminin azalmasıdır12. Bu direnç mekanizmalarının ikisi de kromozomal kaynaklıdır. Ancak ilk kez 1998 yılında bir K.pneumoniae izolatından 218 aminoasitten oluşan tekrarlayan pentapeptid ailesine ait olan ve qnr olarak adlandırılan plazmidle aktarılan yeni bir gen bölgesi saptanmış ve bu gen bölgesi qnrA olarak adlandırılmıştır9. Qnr determinantlarının akuatik bakteri türlerinden, Aeromonas spp., Shewanella algae ve Vibrio splendidus'tan orijin almış olabileceği düşünülmektedir9,13.
Türkiye'de daha önce farklı merkezlerde Enterobacteriaceae ailesinde qnr genleri araştırılmıştır. Ülkemizde 2005 yılında yapılan bir çalışmada, 49 izolatta qnrA gen bölgesi araştırılmış ve bir E.cloacae bir de C.freundii izolatında qnrA geni tespit edilmiştir14. Daha sonra Öktem ve arkadaşları15 tarafından yapılan çalışmada, kan kültürlerinden izole edilen 356 Enterobacteriaceae üyesinde qnrA, qnrB ve qnrS genlerinin varlığı araştırılmış, 61 izolatta qnrA, 3 izolatta qnrS geni saptanmıştır. QnrA saptanan izolatların ikisinin ve qnrS saptanan izolatların birinin GSBL pozitif olduğu gözlenmiştir. Ülkemizde yapılan başka bir çalışmada, yoğun bakım hastalarından izole edilen toplam 460 gram-negatif bakteride qnrA, qnrB, qnrS genleri araştırılmış, 3 (%0.65) E.cloacae izolatının birinde qnrB1 ve ikisinde qnrS1 saptanmıştır16. Türkiye'de farklı hastanelerden izole edilen ve toplam 248 E.coli ve K.pneumoniae izolatında qnrA, qnrB, qnrS, aac(6')-Ib-cr genlerinin sıklığının araştırıldığı bir çalışmada da, bir K.pneumoniae izolatında farklı plazmidler üzerinde qnrB1 ve aac(6')-Ib-cr genleri saptanmıştır17. Mevcut literatür bulgularına göre çalışmamız, Türkiye'de çok merkezli olarak dört qnr gen ailesinin de araştırıldığı ve farklı qnrB24 ve qnrB27 alt tiplerinin de belirlendiği ilk çalışmadır. İspanya'da yapılan çok merkezli bir çalışmada, 19.010 izolat (18.624 Salmonella spp., 285 E.coli, 68 Shigella spp., 29 K.pneumoniae, 2 C.freundii ve 2 Proteus mirabilis) kinolon direnci açısından araştırılmış ve azalmış siprofloksasin duyarlılığı (MİK 0.12- 0.5 mg/L) gösteren ancak nalidiksik aside duyarlı olan 123 izolatta qnr genleri araştırılmış ve iki Salmonella spp. izolatında qnrB, 25 Salmonella spp. ve bir E.coli izolatında qnrA, dört Salmonella spp. izolatında qnrS gen varlığı saptanmıştır7. Yine 13 Avrupa ülkesinden toplanan insan, hayvan, besin ve çevreden izole edilen 485 Salmonella spp. ve 133 E.coli izolatında plazmidle aktarılan kinolon direnç genlerinin varlığı araştırılmış ve Salmonella spp. izolatlarının %59 (288/485)‘unda, E.coli izolatlarının %15 (20/133)'inde pozitiflik saptanmıştır. QnrA üç Salmonella spp. izolatında, qnrB 138 Salmonella spp. ve bir E.coli izolatında, qnrS 125 Salmonella spp. ve 19 E.coli izolatında tespit edilmiştir18.
Qnr gen ailesi alt varyantlarla genişlemekte ve qnrA, qnrB, qnrS ve qnrC genlerinin tüm alt varyantlarının multipleks PCR yöntemiyle tanımlanması güçlük arz etmektedir. Multipleks PCR işlemiyle çalışmamızda da olduğu gibi pozitiflik oranları çok yüksek bulunmuş ve sonrasında tek primer ile pozitif bulunan izolatlar için tekrar PCR yapılmak zorunda kalınmıştır. Tek primerle yapılan PCR ile pozitiflik oranları önemli ölçüde azalmış olup bu izolatlarda qnr varlığı sekanslamayla doğrulanmıştır. Çalışmada çok fazla sayıda izolata yapılan multipleks PCR işleminin zaman, iş yükü ve maliyet açısından çok etkin olmadığı gözlenmiştir. Guillard ve arkadaşları19 yaptıkları çalışmada, qnrA, qnrB, qnrC, qnrS, qnrD ve qepA genlerini gerçek zamanlı PCR yöntemiyle araştırmışlar ve plazmid aracılı kinolon direncinin tespitinde hızlı bir yöntem tanımlamışlardır.
Çalışmamızda, dört farklı merkezden toplanan Enterobacteriaceae ailesine ait 647 klinik izolatta qnr genleri araştırılmıştır. İlk aşamada pozitif suş sayısı çok görülmekle birlikte tekrar tek primerle yapılan PCR işlemi ve sonrasında sekans işlemi sonucunda pozitif izolat sayısı oldukça azalmıştır. Yapılan çalışmalarda qnr genlerinin sıklığının değişkenlik göstermesi, farklı duyarlılık paternine sahip izolatların çalışmalara dahil edilmesi olabilir7,20. Çalışmamızda elde ettiğimiz sonuçlar, qnrA, qnrB ve qnrS gen sıklığının düşük olduğunu göstermektedir. Ayrıca, çalışmamız ülkemizde qnrC geninin araştırıldığı ilk çalışmadır ve çalışmada qnrC pozitifliği saptanmamıştır. Sonuç olarak, günümüzde hızla direnç gelişimi birçok antibakteriyel ajan gibi kinolon kullanımını da sınırlamaktadır. Bu nedenle direnç gelişim yollarının tespit edilip, bunlarla ilgili çalışma sayılarının artırılması ve dünya çapındaki yayılımın belirlenmesi tedavi stratejilerinin geliştirilmesinde yardımcı olacaktır.
TEŞEKKÜR
Çalışmamızda kullanılmak üzere gönderdikleri qnr pozitif suşlar dolayısıyla sayın Prof. G. A. Jacoby, Sayın Prof. P. Nordmann ve Sayın Prof. M. Wang'a teşekkürlerimizi sunarız.
KAYNAKLAR
- Cattoir V, Nordmann P. Plasmid-mediated quinolone resistance in gram-negative bacterial species: an update. Curr Med Chem 2009; 16(8):1028-46. [Özet]
- Ruiz J. Mechanisms of resistance to quinolones: target alterations, decreased accumulation and DNA gyrase protection. J Antimicrob Chemother 2003; 51(5): 1109-17. [Özet] [Tam Metin] [PDF]
- Hooper Dc, Strahilevitz J. Quinolones, pp: 487-510. In: Mandell GL, Bennett JE, Dolin R (eds), Mandell, Douglas and Bennett's Principles and Practice of Infectious Diseases. 2009, 7th ed. Elsevier Churchill Livingstone, Philadelphia.
- Wolfson JS, Hooper D. Fluoroquinlone antimicrobial agents. Clin Microb Rev 1989; 2(4): 378-424. [Özet] [PDF]
- Martinez LM, Pascual A, Jacoby GA. Quinolone resistance from a transferable plasmid. Lancet 1998; 351(9105): 797-9. [Özet]
- Yamane K, Wochino J, Suzuki S, et al. New plasmid-medited quinolone efflux pump, QepA, found in an Escherichia coli clinical isolate. Antimimicrob Agents Chemother 2007; 51(9): 3354-60. [Özet] [Tam Metin] [PDF]
- Herrera-Leon S, Gonzalez-Sanz R, Herrera-Leon L, Echeita MA. Characterization of multidrug-resistant Enterobacteriaceae carrying plasmid-mediated quinolone resistance mechanisms in Spain. J Antimicrob Chemother 2011; 66(2): 287-90. [Özet] [Tam Metin] [PDF]
- Garcia-Fulgueiras V, Bado I, Mota MI, et al. Extended-spectrum b-lactamases and plasmid-mediated quinolone resistance in enterobacterial clinical isolates in the pediatric hospital of Uruguay. J Antimicrob Chemother 2011; 66(8): 1725-9. [Özet]
-
Cattoir V, Poirel L, Aubert C, Soussy CJ, Nordmann P. Unexpected
occurence of plasmid-mediated quinolone resistance determinants in enviromental
Aeromonas spp. Emerg Infect Dis 2008; 14(2): 231-7.
[Özet] [Tam Metin] [PDF] - Strahilevitz J, Jacoby GA, Hooper D, Robicsek A. Plasmid-mediated quinolone resistance: a multifaceted threat. Clin Microb Rev 2009; 22(4): 664-89. [Özet] [Tam Metin] [PDF]
- Kim HB, Park CH, Kim CJ, Kim EC, Jacoby GA, Hooper DC. Prevalence of plasmid-mediated quinolone resistance determinants over a 9-year period. Antimicrob Agents Chemother 2009; 53(2): 639-45. [Özet] [Tam Metin] [PDF]
-
Jacoby GA. Mechanisms of resistance to quinolones. Clin Infect Dis 2005;
41(Suppl 2): S120-6.
[Özet] [Tam Metin] [PDF] - Poirel L, Rodriguez-Martinez JM, Mammeri H, Liard A, Nordmann P. Origin of plasmid-mediated quinolone resistance determinant QnrA. Antimicrob Agents Chemother 2005; 49(8):3523-5. [Özet] [Tam Metin] [PDF]
- Nazik H, Öngen B. Türkiye'de plazmid aracılı kinolon direnci. ANKEM 2010; 24(1): 46-54. [Özet] [PDF]
- Öktem MA, Biçmen M, Gülay Z. Kan kültürlerinden soyutlanan Enterobacteriaceae izolatlarında plazmid ile ilişkili kinolon direnci genlerinin saptanması. 8. Antimikrobik Kemoterapi Günleri, 2-4 Nisan 2008, İstanbul. Program ve Özet Kitabı, P28.
- Nazik H, Ongen B, Kuvat N. Investigation of plasmid-mediated quinolone resistance among isolates obtained in a Turkish intensive care unit. Jpn J Infect Dis 2008; 61(4): 310-2. [Özet] [PDF]
- Poirel L, Gür D, Minarini L, Arslan U, Nordmann P. Molecular epidemiology of plasmid-mediated quinolone resistance determinants in extended spectrum beta-lactamase producing E.coli and K.pneumoniae isolates from Turkey. 18th European Congress of Clinical Microbiology and Infectious Diseases, 19-22 April 2008, Barcelona. Abstract Book, P1527.
- Veldman K, Cavaco LM, Mevius D, et al. International collaborative study on the occurence of plasmid-mediated quinolone resistance in Salmonella enterica and Escherichia coli isolated from animals, humans, food and the enviroment in 13 European countries. J Antimicrob Chemother 2011; 66(6): 1278-86. [Özet] [Tam Metin] [PDF]
- Guillard T, Moret H, Brasme L, et al. Rapid detecion of qnr and qepA plasmid-mediated quinolone resistance genes using real-time PCR. Diagn Microbiol Infect Dis 2011; 70(2):253-9. [Özet]
- Park YJ, Yu JK, Kim SY, Lee S, Jeong SH. Prevalence and characteristics of qnr determinants and aac(6')-Ib-cr among ciprofloxacin-susceptible isolates of Klebsiella pneumoniae in Korea. J Antimicrob Chemother 2010; 65(9): 2041-3. [Tam Metin] [PDF]
İletişim (Correspondence):
Doç. Dr. Ahmet Yılmaz Çoban,
Ondokuz Mayıs Üniversitesi Tıp Fakültesi,
Tıbbi Mikrobiyoloji Anabilim Dalı,
Samsun, Türkiye.
Tel (Phone): +90 362 457 6070,
E-posta (E-mail): cobanay2003@yahoo.com.tr