İnsan Papillomavirusunun Genomik Yapısı ve Proteinleri
Genomic Organization and Proteins of Human Papillomavirus
Gülçin ALP AVCI
Gazi
Üniversitesi Tıp Fakültesi, Tıbbi Mikrobiyoloji Anabilim Dalı, Ankara - Hitit
Üniversitesi Sağlık Yüksekokulu,
Mikrobiyoloji Bölümü, Çorum.
Gazi University Faculty of Medicine, Department of Medical Microbiology, Ankara - Hitit University School of Health, Department of Microbiology, Corum, Turkey.
ÖZET
İnsan papillomavirus (HPV) enfeksiyonları, dünyada cinsel yolla bulaşan hastalıkların en önemlilerinden biridir. Bugün için, DNA dizisi kullanılarak tanımlanan 200'den fazla HPV tipi bulunmaktadır. HPV tipleri onkojenik potansiyellerine göre; yüksek riskli (tip 16, 18, 31, 33, 35, 39, 45 vb.), olası yüksek riskli (tip 26, 53, 66) ve düşük riskli (tip 6, 11, 40, 42, 43, 44, 54 vb.) tipler olarak gruplandırılmaktadır. Yüksek riskli HPV tipleri, günümüzde servikal intraepitelyal neoplazi ve serviks kanserinin temel etiyolojik faktörü olarak kabul edilmektedir. Papillomaviridae ailesinde yer alan HPV tipleri, çıplak, ikozahedral simetrili, yaklaşık 55 nm çapında viruslardır. Çift iplikli çembersel DNA'dan oluşan 8 kb büyüklüğündeki genom; virusun replikasyonunda ve hücre transformasyonunda rol oynayan erken proteinler (E1, E2, E4, E5, E6, E7) ile kapsid yapısını oluşturan geç proteinleri (L1, L2) kodlar. HPV DNA'sının konak hücre kromozomuna integrasyonu, virusun kalıcılığı ve karsinojenik etkileri için önem taşımaktadır. İntegrasyon genom boyunca rastgele meydana gelebilir ve bu durum virusun onkoproteinleri olan E6/E7 gen ürünlerinin ekspresyonunda artışa, dolayısıyla da onkogeneze yol açabilir. Ancak integrasyon olmadan da E6/E7 ekspresyon artışı gerçekleşebilmektedir. Servikal kanser hücrelerinin çoğunda HPV integrasyonu genellikle E2 gen bölgesinde kırılmaya neden olmakta ve bu durum E6 ve E7 onkogenlerinin ekspresyonunda artışa yol açmaktadır. Özellikle HPV-16 ve HPV-18 gibi yüksek riskli tiplerin E6 ve E7 proteinleri, sırasıyla p53 ve retinoblastoma proteini (pRb) gibi tümör baskılayıcı proteinler ile etkileşip onların fonksiyonlarını inhibe etmekte; böylece hücre çoğalmasının kontrolünü bozmakta ve hücrelerin ölümsüzleşmesine neden olmaktadır. E6 proteininin p53'e bağlanması hücrenin G1 fazında tutulmasına, apoptoz ve DNA tamirinin durdurulmasına yol açar. E7 proteini ise, pRb ve siklin-E gibi mitotik interaktif hücresel proteinler ile ilişkiye girerek, hücresel DNA sentezi ve hücre çoğalmasının uyarılmasında rol oynar. Son yıllarda tanımlanan ve sadece bazı papillomavirus tiplerinde (HPV 1, 11, 16, 31, 33) bulunan E3 ve E8 genleri de erken gen bölgesinde yer almaktadır. Bir füzyon proteini olan E8^E2C'nin, HPV DNA replikasyonunun negatif bir regülatörü olduğu ve viral kopya sayısını kontrol ederek HPV epizomlarının stabilitesini sağladığı düşünülmektedir. Bu derleme yazıda HPV'nin genom yapısı ve gen ürünlerinin fonksiyonları özetlenmektedir.
Anahtar sözcükler: İnsan papillomavirusu; genom yapısı; viral protein; E2; E6; E7.
ABSTRACT
Human papillomavirus (HPV) infections are one of the most common sexually-transmitted diseases worldwide. Nowadays, more than 200 HPV types have been identified by DNA sequencing. HPV types are also grouped into three, such as high-risk (types 6, 11, 40, 42, 43, 44, 54, etc), probable high-risk (types 26, 53, 66) and low-risk (types 6, 11, 40, 42, 43, 44, 54, etc) types according to their oncogenic potential. HPV is currently considered as the main aetiological factor of cervical intraepithelial neoplasia and cervical cancer. HPV types classified in Papillomaviridae family, are non-enveloped, icosahedral symmetric viruses about 55 nm in size. Viral genome consists of circular double-stranded DNA, about 8 kb in size, encodes for early proteins (E1, E2, E4, E5, E6, E7) which play role in virus replication and cell transformation, and for late (L1, L2) proteins which are the structural units of the viral capsid. Integration of HPV DNA into the host chromosome is crucial for viral persistence and for carcinogenic effects. Viral DNA may integrate randomly to the cell genome and integration can lead to the deregulation and increase of E6/E7 expression leading to oncogenesis. However, increased expression of E6/E7 gene products may occur without genome integration. E6 and E7 proteins of especially high-risk HPV types (e.g. types 16 and 18) interact with tumor supressor proteins such as p53 and retinoblastoma (pRb) proteins, respectively; inhibit their functions and cause uncontrolled proliferation and immortalization of the cells. The binding of E6 protein to p53 leads its rapid degradation, and the eclipse in the G1 phase, DNA repair mechanisms and apoptosis are terminated. In the other way, E7 protein interacts with pRb and mitotically interactive cellular proteins such as cyclin-E, causing stimulation of cellular DNA synthesis and cell proliferation. Recently identified genes E3 and E8 are located in early gene region and found only in a few papillomavirus types (HPV 1, 11, 16, 31, 33). A fusion protein, E8^E2C, functions as a negative regulator for HPV DNA replication and it is thought that this protein may play a role in the control of viral copy number as well as in the stable maintenance of HPV episomes. In this review article, the genomic structure of HPV and the functions of gene products have been summarized.
Key words: Human papillomavirus; genomic organization; viral protein; E2; E6; E7.
Geliş Tarihi (Received): 14.12.2011 • Kabul Ediliş Tarihi (Accepted): 06.04.2012
GİRİŞ
Papillomavirusların onkojenik potansiyeli ile ilgili uzun yıllar süren araştırma ve tartışmaların sonunda, insan papillomavirusları ile servikal kanser arasındaki ilişkinin 1976 yılında Harald zur Hausen tarafından gösterilmesi, bu virusların virolojik ve klinik önemini daha da artırmıştır1. Bugün için insan papillomaviruslarının yüksek riskli tipleri, tüm dünyada serviks kanserinin primer etiyolojik ajanı olarak kabul edilmekte; özellikle tip 16 ve 18 servikal kanserlerin %70'inden sorumlu tutulmaktadır2,3.
İnsan papillomavirusları (Human papillomavirus; HPV), Papillomaviridae ailesinde sınıflandırılan, çıplak, ikozahedral simetrili, yaklaşık 55 nm çapında, çift iplikli DNA içeren viruslardır. Viral kapsid 60 hekzon ve 12 penton olmak üzere toplam 72 kapsomerden oluşur; protein kılıf, bir majör ve bir minör olmak üzere iki kapsid proteini içerir4. Bu viruslar DNA yapılarına göre sınıflandırıldığından serotipler yerine genotipler olarak ve keşfedildikleri sıraya göre numaralandırılır4,5. HPV'lerin sınıflandırılması, tür orijini ve DNA hibridizasyonuyla tespit edilen viral genomlar arasındaki homolojinin derecesine bağlıdır. Günümüzde 200'den fazla HPV tipi tanımlanmıştır. HPV tipleri ayrıca, klinik olarak karsinojenik potansiyellerine göre de; (1) düşük riskli HPV tipleri (6, 11, 40, 42, 43, 44, 54, 61, 70, 72, 81 ve CP6108), (2) olası yüksek riskli HPV tipleri (26, 53 ve 66) ve (3) yüksek riskli HPV tipleri (16 başta olmak üzere 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 68, 73 ve 82) olmak üzere üç gruba ayrılmışlardır4,6.
İnsan papillomavirusları dünyada yaygın olarak bulunmakta ve bulaşma direkt temas ya da cinsel yolla olmaktadır7,8. Virus insanda primer olarak skuamöz epitel hücreleri ve mukozayı enfekte etmekte; deri ve mukozalarda bazal tabakalardan başlayarak çeşitli lezyonlar oluşturmaktadır. Düşük riskli HPV tipleri (özellikle HPV-6 ve HPV-11) genital siğiller ve rekürrent larengeal papillomatoz gibi benign lezyonlara neden olurken, yüksek riskli HPV tipleri servikal, vajinal, vulvar, anal, penil ve orofarengeal karsinomayı da içine alan çeşitli kanserlerin gelişiminde doğrudan ya da dolaylı olarak rol oynayabilmektedir2,6,7,8,9. HPV enfeksiyonlarının sonucu, konak faktörlerine bağlı olduğu gibi virusun tipine ve enfekte olan bölgeye de bağlıdır.
HPV GENOMİK YAPISI
Papillomaviruslar, küçük boyutlarına rağmen moleküler biyolojileri oldukça kompleks olan viruslardır. Çift iplikli çembersel DNA'dan oluşan genom yaklaşık 8 kilobaz büyüklüğündedir ve viral proteinleri kodlayan diziler tek bir DNA sarmalı üzerindedir5. Fonksiyonel olarak erken bölge (early; E), geç bölge (late; L) ve uzun kontrol bölgesi (long control region; LCR) olmak üzere üç bölgeye ayrılır. Tüm papillomaviruslarda bu üç bölge, erken pA ve geç pA olarak iki poliadenilasyon (pA) bölgesi tarafından ayrılmaktadır. Erken bölgede sekiz, geç bölgede ise iki açık okuma çerçevesi (Open reading frame; ORF) bulunur. Bütün HPV ORF'ları virusun sadece bir sarmalı üzerinde yer alır ve 10 ORF viral replikasyon döngüsündeki gen ekspresyon sırasına göre erken ve geç olarak isimlendirilir. Erken proteinler (E1, E2, E4, E5, E6 ve E7) viral replikasyon ve hücre transformasyonunda rol oynar5,10. Son yıllarda keşfedilen E3 ve E8'de aynı bölgede bulunmakta olup, E8'in E2 bölgesinin delesyonu sırasında ortaya çıktığı düşünülmektedir11. Geç gen bölgesinde ise L1 (majör kapsid proteini) ve L2 (minör kapsid proteini) proteinleri kodlanmaktadır (Şekil 1)12. Tüm bu proteinler, transmembranın uyarılması, hücre döngüsünün düzenlenmesi ve transformasyon aktivitesinin denetlenmesi gibi pleotropik fonksiyonlara sahiptir12.
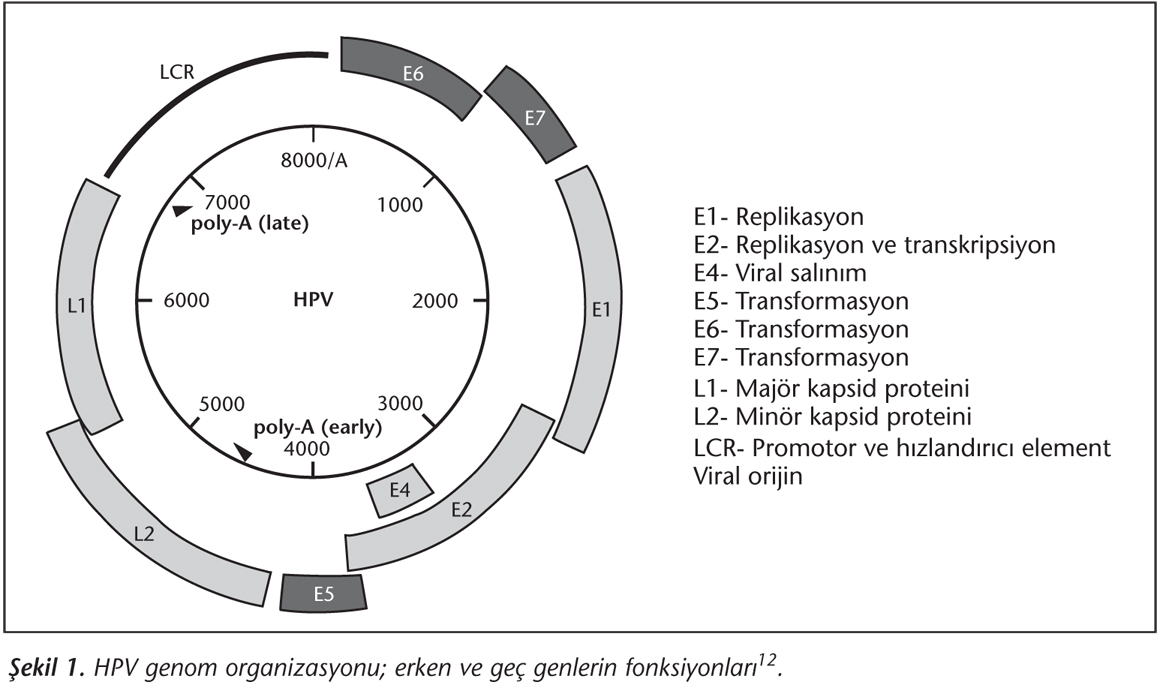
HPV PROTEİNLERİ
E1 ve E2 Proteinleri
E1 ve E2 genleri HPV'nin önemli düzenleyici proteinlerini kodlamaktadır. Bu genler parabazal keratinositlerde bulunur; viral epizomun dengeye getirilmesinde, viral replikasyonun başlatılmasında ve replikasyonda rol oynar. E2 tarafından kodlanan protein LCR'deki E2 bağlanma yerine bağlanarak transkripsiyonu baskılar. E1 tarafından kodlanan protein ise promoter bölgesine E2'nin bağlanmasını kolaylaştırır13,14. E2 proteini, prodüktif enfeksiyon sırasında çeşitli roller üstlenmektedir. Bu protein, bazal hücrelerde viral DNA replikasyonunun başlaması ve genomun açılması için gereklidir14. Farklılaşmamış hücrelerde E2 proteini, düşük dozlarda transkripsiyonu artırmak için bir viral transkripsiyonel transaktivatör gibi davranırken, yüksek konsantrasyonlarda transkripsiyonel faktörlerin, onları tanıyan dizilere bağlanmasını engelleyerek baskılayıcı hale gelebilir15. Farklılaşmış hücrelerde ise E2 proteini, geç promoter bölgesindeki değişimden dolayı transkripsiyonda baskılayıcı fonksiyonunu kaybeder. Bu durum, E1 ve E2 protein seviyelerinin ve bu sebeple viral DNA amplifikasyonunun hızlı artışıyla sonuçlanır. Servikal kanser hücrelerinin çoğunda, HPV integrasyonu genellikle E2 gen bölgesinde kırılmaya neden olur ve bu kırılma E2 proteininin fonksiyon kaybıyla sonuçlanır15,16. Aynı zamanda elde edilen veriler, E2 gen bölgesinde meydana gelen bu kırılmayla birlikte, P97 promotor aktivitesinde ve E6 ile E7 viral onkogenlerin ekspresyonunda artış olduğunu göstermektedir16. E2 aynı zamanda viral genom replikasyonu için de gereklidir. E2 özgül olarak E1'i tanır ve viral replikasyon orijininde (Ori) E1'i toplar, burada E1-E2 kompleksi oluşur17. Yapılan çalışmalarda ayrıca, E2 aktivasyonunun, HPV ile enfekte hücrelerde apoptozu ve yaşlanmayı uyardığı, buna karşın E6 ve E7 transkripsiyonunu kısmen inhibe ettiği belirlenmiştir10,13,18.
E1 proteini, 68 kDa'luk ATPase, DNA helikaz ve DNA sarmalının açılması için aktiviteye sahip olan nükleer bir proteindir. Düşük kuvvetle viral Ori'ye bağlanır ve bağlanma afinitesi E2 proteini ile artırılabilir. E1, DNA replikasyonunu aktive etmek için DNA polimeraz-a'nın alt üniteleriyle etkileşir. E1 proteini, hem viral DNA replikasyonunun başlamasında hem de replikasyon çatalında helikaz aktivitesine bağlı olarak elongasyonda rol oynamaktadır17.
E4 Proteini
E4 proteini enfeksiyonun geç döneminde bol miktarda üretilir. Tam uzunlukta bir E4 proteini, uç uca eklenmiş E1-E4 transkriptlerinden oluşur. E4 ORF ile E1'in ilk beş aminoasidi 17 kDa'luk E1-E4 füzyon proteinini oluşturur. E1-E4 transkriptleri HPV'nin replikasyon döngüsü süresince salınmaktadır. Tam uzunluktaki E1-E4 proteini, üst farklılaşmış hücre tabakalarında proteolitik olarak parçalanmış polipeptid ürünlerine bağlıdır ve en yüksek düzeyde salınım, farklılaşmış suprabazal tabakada meydana gelir13,15. Virusun hayat döngüsünde E1-E4'ün fonksiyonu hala tam olarak açık değildir. E1-E4 proteini özellikle sitoplazmada yer almaktadır. Yüksek riskli HPV tiplerinin E4 proteini, enfekte hücrelerin sitoplazmasında yaygın olarak dağılır ve keratin aracı (intermediate) filamentleri (IF) ile organize filamentöz bir ağ formu oluşturur. Ayrıca in vitro deneyler, bu proteinin hücrenin iskelet yapısında bozulmalara (sitoskeletal kollaps) yol açarak virusun salınımını kolaylaştırdığını düşündürmektedir17,19.
E5 Proteini
E5 proteini yaklaşık 5 kDa büyüklüğünde, 83 aminoasitlik küçük bir hidrofobik membran proteinidir. Başlıca endoplazmik retikulumda, Golgi cisimciğinde ve plazma membranlarında bulunmaktadır. E5 proteini, enfeksiyonun başlangıcında oldukça önemlidir. Epidermal büyüme faktörü reseptörü (EGFR), trombosit aktive edici faktör reseptörü (PAFR) ve koloni uyarıcı faktör (CSF)-1 reseptörü ile bir kompleks oluşturarak hücre çoğalmasını uyarır14. Ayrıca E5 proteini, apoptozu izleyen DNA hasarında da rol oynamaktadır. HPV enfeksiyonu ile gelişen servikal kanser lezyonlarında, epizomal viral DNA sıklıkla konak DNA'sına entegre olmakta ve E5 proteinini kodlayan diziyi de içeren genomun azımsanmayacak bir parçası delesyona uğramaktadır. Bu nedenle, HPV ile ilişkili karsinogenezin geç dönemlerinde E5 zorunlu değildir13,14. Son zamanlarda yapılan çalışmalar, E5 proteininin, virusun çoğalması ve farklılaşmış hücrelerde viral gen ekspresyonunun modülasyonunda E7 ile ortak bir rol oynadığını göstermiştir15.
E6 ve E7 Proteinleri
HPV'lerin onkojenik fonksiyonları, özellikle E6 ve E7 proteinlerinin üretimine bağlıdır; zira E6 ve E7, HPV'nin iki temel onkoproteinidir. Çeşitli çalışmalarda, HPV E6 ve E7 genlerinin tümör süpresör genlerinin fonksiyonlarını baskılamadaki kritik rolü vurgulanmaktadır2,6,13,17. E6 onkoproteini, yaklaşık 17 kDa büyüklüğünde olup, yüksek riskli HPV'lerde konak hücrenin hem nükleus hem de sitoplazmasında bulunur. E6 proteini, p53 ve hücresel ubikütinasyon enzim E6-AP'yi (protein yıkımında kullanılan bir enzim) kapsayan üçlü bir kompleksin oluşumuyla, tümör baskılayıcı p53 proteininin parçalanmasını uyararak hücre proliferasyonunu teşvik eder (Şekil 2). E6 ile aktive edilen parçalanma sonucunda, hücre döngüsünün ilerlemesi bozularak tümör hücre gelişiminde artma gerçekleşir20. Bir tümör baskılayıcı protein olan p53'ün, hücre bölünmesi üzerinde negatif regülatör etkisi vardır. Bu etkiyi, hücre döngüsünün G0 ve G2 fazından S fazına geçişini kontrol ederek gerçekleştirmektedir21. Ayrıca p53, hücre döngüsünü durduran veya apoptozu uyaran bir tümör baskılayıcısıdır. Tüm bu fonksiyonlarıyla p53 hücrede meydana gelen DNA hasarlarını gidermek üzere hücre döngüsünü durdurarak, DNA tamiri için gerekli olan onarımın yapılmasını sağlar veya ciddi kromozomal defektleri olan hücrelerde apoptozu indükler. E6, p53 üzerindeki etkisiyle kromozomal onarım yapılmadan hücrelerin bölünmesine, kromozomal defektli hücrelerin birikmesine ve apoptotik etkinin ortadan kalkmasına katkı sağlamaktadır22.
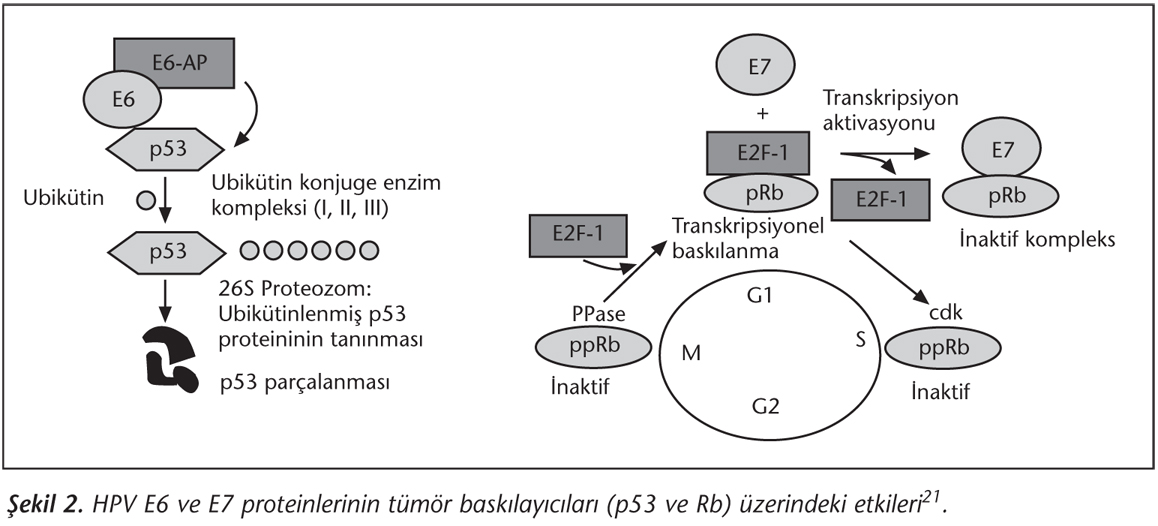
E6'nın apoptotik etkiyi ortadan kaldırması, telomeraz enzimini indükleyerek de gerçekleşmektedir. Telomeraz, kromozomların telomer ucuna hekzanükleotid tekrarları ekleyen bir enzimdir. Telomeraz aktivitesi genellikle embriyonik hücrelerle sınırlıdır ve normal somatik hücrelerde yoktur. Telomeraz olmadığı zaman hücrelerin defalarca bölünmesi, yaşlanması ve fonksiyonlarını kaybetmesi söz konusudur23. Bu enzim, erozyona uğrayan ve kısalan telomerleri tamir ederek otoregülasyonu inhibe eder. Böylece hücrenin yaşamını uzatır ve bu durum apoptozun ortadan kaldırılmasını sağlar24. Ancak HPV E6 proteini, telomeraz ekspresyonunu indüklemektedir15.
E7 onkoproteini ise, yaklaşık 12 kDa molekül ağırlığındadır ve öncelikle nükleusta bulunur. Yüksek riskli HPV E7 proteinlerinin temel aktivitesi, retinoblastom (Rb) ailesi üyeleriyle ilişkisi sebebiyle hücrelerin farklılaşmasında rol oynamasıdır. Düşük riskli HPV'nin E7 proteinlerinin aynı zamanda Rb'ye düşük kuvvetle bağlandığı da gösterilmiştir19. Rb proteini DNA hasarına bağlı olarak hücre döngüsünü durdurur veya apoptozu indükler24. E7 proteini Rb'ye cep kangalı (domain) adı verilen bir bölgeden bağlanır. Rb'nin cep kangalı dizisi, tümör baskılayıcı fonksiyonu için önemlidir. Rb proteini, hücre döngüsünün G1 fazından S fazına geçişini inhibe eder ve aktivitesi fosforilasyonla düzenlenir. HPV ile enfekte hücrelerde, E7 proteinlerinin hipofosforile Rb proteinlere bağlanması, Rb proteininin inaktivasyonuyla sonuçlanır ve hücre döngüsünün S fazına ilerlemesine izin verir19,20,25. Rb'nin önemli biyokimyasal fonksiyonlarından biri de, E2F transkripsiyon faktörlerine bağlanmak ve replikasyonda görevli olan enzim genlerinin aktivasyonunu baskılamaktır. Replikasyonda görevli enzim genlerinin baskılanma yeteneği, Rb'nin tümör baskılayıcı fonksiyonuyla ilişkilidir22,25,26. E7'nin Rb proteini (pRb) ve cep protein ailesinin diğer üyeleriyle etkileşimi, büyüme faktörlerinin varlığından bağımsız olarak, pRb ve transkripsiyon faktörlerinin E2F ailesi arasındaki ilişkiyi bozmaktadır. E7 aynı zamanda histon deasetilazları, AP1 transkripsiyon kompleksini ve siklin-bağımlı kinaz (SBK) inhibitörleri p21 ve p27 bileşenlerini içeren hücre proliferasyonuyla ilgili diğer proteinlerle de etkileşmektedir25. Doğal enfeksiyon sırasında ise, p21 ve p27 SBK inhibitörlerinin düzeylerine bağlı olarak bazı hücreler inhibe olur; buna karşın E7 hücre çoğalmasının sürekliliğini sağlar14.
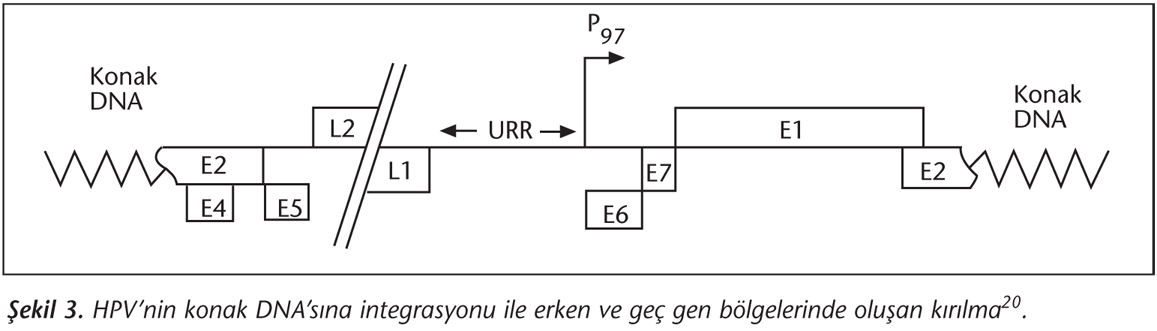
Kanserin gelişiminde E2 bölgesinde sıklıkla kırılma meydana geldiğinden HPV DNA integrasyonu önemlidir (Şekil 3)20. İntegrasyon esnasında E2'de meydana gelen kırılma, E2'nin E6 ve E7 üzerindeki inhibitör etkisini ortadan kaldırır ve E8 bölgesinin oluşumuna yol açar. Sonuçta, E6 ve E7 viral proteinlerinin kontrolsüz olarak eksprese edildiği görülür19,20. Bu nedenle, E6 ve E7 onkoproteinleri kanser gelişimi için önemlidir.
E8 Proteini
E8 proteininin açık okuma bölgesi, sığır papillomavirus tip 1 (BPV-1) ve HPV tip 31 (HPV-31) dışında, birçok tavşan papillomaviruslarında bir onkogen olarak tespit edilmiştir. Ayrıca E8^E2 füzyon proteini, BPV-1 ve HPV-31'de viral transkripsiyon ve replikasyonun negatif bir düzenleyicisi olarak belirlenmiştir26,27. E8 geninin bir parçası olan E8^E2 füzyon proteini, E2 geninin C terminaline bağlanmış olup, viral transkripsiyonun alternatif bir yolunu oluşturmaktadır. E8^E2C'nin, viral yaşam döngüsünün erken evresi boyunca HPV DNA replikasyonunu sınırlandırdığı ve viral kopya sayısını kontrol ederek HPV epizomlarının stabilitesini sağladığı düşünülmektedir27. Yapılan bir çalışmada, E8^E2C eksprese edemeyen HPV-31 mutantlarının, yüksek düzey DNA replikasyonuna rağmen insan keratinositlerinde ekstrakromozomal olarak kalamadığı gösterilmiştir28.
L1 ve L2 Proteinleri
Viral DNA'nın geç bölgesi yaklaşık 3 kb uzunluğunda olup, sırasıyla majör ve minör proteinleri kodlayan L1 ve L2 genlerini içermektedir. L1 proteini yaklaşık 55 kDa, L2 proteini ise yaklaşık 74 kDa büyüklüğünde olan kapsid proteinleridir. L1 proteini, hücreye giriş için a6b4 integrin, heparan sülfat ve glikozaminoglikan dahil üç farklı yüzey reseptörüne bağlanır. L2'nin hücresel yüzey proteinine bağlanması da, HPV enfeksiyonu için gereklidir19. Enfekte hücrenin sitoplazmasında L2 proteini sitoplazmadan nükleusa virusun etkili transportu için beta-aktin ile etkileşir. L1 ve L2 proteinlerinin reseptör aracılı bir yolla nükleus içine taşınması önemlidir. L2 proteininin DNA'ya bağlanmasından ve enkapsidasyon olaylarından sorumlu olduğu ileri sürülmektedir29. Son zamanlarda HPV-16 L2 proteini varlığının, E2'nin transkripsiyonel transaktivasyonunu inhibe ettiği, ancak DNA replikasyonunu etkilemediği bildirilmiştir30. Viral hayat döngüsünün düzenlenmesinde L2-E2 birleşmesinin etkisi hala araştırılmaktadır13,15,17.
KAYNAKLAR
- zur Hausen H. Papillomaviruses in the causation of human cancers-a brief historical account. Virology 2009; 384(2): 260-5. [Özet]
- Münger K, Baldwin A, Edwards KM, Hayakawa H, Nguyen CL, Owens M. Mechanisms of human papillomavirus-induced oncogenesis. J Virol 2004; 78(21): 11451-60. [Tam Metin] [PDF]
-
Lowy DR, Schiller JT. Reducing HPV-associated cancer globally. Cancer
Prev Res (Phila) 2012; 5(1): 18-23.
[Özet] [Tam Metin] [PDF] - De Villiers EM, Fauquet C, Broker TR, Bernard HU, zur Hausen H. Classification of papillomaviruses. J Virol 2004; 324(1): 17-27. [Özet]
- zur Hausen H. Papillomaviruses and cancer: from basic studies to clinical application. Nat Rev Cancer 2002; 2(5): 342-50. [Özet]
- No JH, Kim MK, Jeon YT, Kim YB, Song YS. Human papillomavirus vaccine: widening the scope for cancer prevention. Mol Carcinog 2011; 50(4): 244-53. [Özet]
- Yarkın F, Vardar MA. HPV immunolojisi ve natürel enfeksiyonlar. Türkiye Klinikleri J Gynecol Obst Special Topics 2009; 2(1): 43-7. [Özet]
- Srivastava S, Gupta S, Roy JG. High prevalence of oncogenic HPV-16 in cervical smears of asymptomatic women of eastern Uttar Pradesh, India: a population-based study. J Biosci 2012; 37(1): 63-72. [Özet] [PDF]
-
Onan MA, Taskiran C, Bozdayi G, et al. Assessment of human papilloma
viral load of archival cervical intraepithelial neoplasia by real-time
polymerase chain reaction in a Turkish population. Eur J Gynaecol Oncol 2005;
26(6): 632-5.
[Özet] - Leto MG, Santos Júnior GF, Porro AM, Tomimori J. Human papillomavirus infection: etiopathogenesis, molecular biology and clinical manifestations. An Bras Dermatol 2011; 86(2): 306-17. [Özet]
- Motoyama S, Cecilia A. Ladines-Llave, Villanueva SL, Maruo T. The role of human papillomavirus in the molecular biology of cervical carcinogenesis. Kobe J Med Sci 2004; 50(1): 9-19. [Özet] [PDF]
- Howley PM. The molecular biology of papillomavirus transformation. Am J Pathol 1983; 113(3): 414-21. [PDF]
- Hebner CM, Laimins LA. Human papillomaviruses: basic mechanisms of pathogenesis and oncogenicity. Rev Med Virol 2006; 16(2): 83-97. [Özet]
- Doorbar J. Molecular biology of human papillomavirus infection and cervical cancer. Clin Sci (Lond) 2006; 110(5): 525-41. [Özet] [Tam Metin]
- Longworth MS, Laimins LA. Pathogenesis of human papillomavirus in differentiating epithelia. Microbiol Mol Biol Rev 2004; 68(2): 362-72. [Özet] [Tam Metin] [PDF]
- Tsakogiannis D, Ruether IG, Kyriakopoulou Z, et al. Sequence variation analysis of the E2 gene of human papilloma virus type 16 in cervical lesions from women in Greece. Arch Virol 2012; DOI 10.1007/s00705-012-1236-8. [Özet]
- Thomison J 3rd, Thomas LK, Shroyer KR. Human papillomavirus: molecular and cytologic/histologic aspects related to cervical intraepithelial neoplasia and carcinoma. Hum Pathol 2008; 39(2): 154-66. [Özet]
- Kadaja M, Isok-Paas H, Laos T, Ustav E, Ustav M. Mechanism of genomic instability in cells infected with the high-risk human papillomaviruses. PLoS Pathog 2009; 5(4): e1000397. [Özet] [Tam Metin] [PDF]
-
Burd EM. Human papillomavirus and cervical cancer. Clin Microbiol Rev
2003; 16(1): 1-17.
[Özet] [Tam Metin] [PDF] - Fehrmann F, Laimins LA. Human papillomaviruses: targeting differentiating epithelial cells for malignant transformation. Oncogene 2003; 22(33): 5201-7. [Özet] [Tam Metin] [PDF]
- Yim EK, Park JS. The role of HPV E6 and E7 oncoproteins in HPV-associated cervical carcinogenesis. Cancer Res Treat 2005; 37(6): 319-24. [Özet] [Tam Metin] [PDF]
- Hubbert NL, Sedman SA, Schiller JT. Human papillomavirus type 16 E6 increases the degradation rate of p53 in human keratinocytes. J Virol 1992; 66(10): 6237-41. [Özet] [PDF]
- Köse F, Turan T. Servikal kanser tümörogenezi ve HPV. Türkiye Klinikleri J Gynecol Obst Special Topics 2009; 2(1): 13-8. [Özet]
- Kiyono T, Foster SA, Koop JI, et al. Both Rb/p16INK4a inactivation and telomerase activity are required to immortalize human epithelial cells. Nature 1998; 396(6706): 84-8. [Özet]
- Ganguly N, Parihar SP. Human papillomavirus E6 and E7 oncoproteins as risk factors for tumorigenesis. J Biosci 2009; 34(1): 113-23. [Özet] [PDF]
- Munoz N, Bosch FX, de Sanjose S, et al. Epidemiologic classification of human papillomavirus types associated with cervical cancer. N Engl J Med 2003; 348(6): 518-27. [Özet] [Tam Metin] [PDF]
-
Fertey J, Ammermann I, Winkler M, Stöger R, Iftner T, Stubenrauch F.
Interaction of the papillomavirus E8--E2C protein with the cellular CHD6
protein contributes to transcriptional repression. J Virol 2010; 84(18):
9505-15.
[Özet] [Tam Metin] [PDF] - Stubenrauch F, Hummel M, Iftner T, Laimins LA. The E8E2C protein, a negative regulator of viral transcription and replication, is required for extrachromosomal maintenance of human papillomavirus type 31 in keratinocytes. J Virol 2000; 74(3): 1178-86. [Özet] [Tam Metin] [PDF]
- Finnen RL, Erickson KD, Chen XS, Garcea RL. Interactions between papillomavirus L1 and L2 capsid proteins. J Virol 2003; 77(8): 4818-26. [Özet] [Tam Metin] [PDF]
- Pereira R, Hitzeroth II, Rybicki EP. Insights into the role and function of L2, the minor capsid protein of papillomaviruses. Arch Virol 2009; 154(2): 187-97. [Özet]
İletişim (Correspondence):
Öğr. Gör. Dr. Gülçin Alp Avcı,
Gazi Üniversitesi Tıp Fakültesi
Tıbbi Mikrobiyoloji Anabilim Dalı,
Beşevler, Ankara, Türkiye.
Tel (Phone): +90 312 202 4628
Hitit Üniversitesi Sağlık Yüksekokulu,
Mikrobiyoloji Bölümü,
19030, Çorum, Türkiye.
Tel (Phone): +90 364 223 0730-3509,
E-posta (E-mail): alp.gulcin@yahoo.com