Denizli B�lgesinde Kestane (Castanea sativa) Ağacından Cryptococcus neoformans İzolasyonu
Isolation of Cryptococcus neoformans from a Chesnut Tree (Castanea sativa), Denizli, Turkey
Mustafa ŞENG�L1, Murat KUTLU2, Aylin D�ĞEN3, Levent AKSOY1, Serpil GONCA4, Macit İLKİT5, �ağrı ERGİN1
1 Pamukkale �niversitesi Tıp Fak�ltesi, Tıbbi Mikrobiyoloji Anabilim Dalı, Denizli.
1 Pamukkale University Faculty of Medicine, Department of Medical Microbiology, Denizli, Turkey.
2 Pamukkale �niversitesi Tıp Fak�ltesi, Enfeksiyon Hastalıkları ve Klinik Mikrobiyoloji Anabilim Dalı, Denizli.
2 Pamukkale University Faculty of Medicine, Department of Clinical Microbiology and Infectious Diseases, Denizli, Turkey.
3 Mersin �niversitesi Eczacılık Fak�ltesi, Farmas�tik Mikrobiyoloji Anabilim Dalı, Mersin.
3 Mersin University Faculty of Pharmacy, Department of Pharmaceutical Microbiology, Mersin, Turkey.
4 Mersin �niversitesi, İleri Eğitim, Araştırma ve Uygulama Merkezi, Mersin.
4 Mersin University, Advanced Technology Education Research and Application Center, Mersin, Turkey.
5 �ukurova �niversitesi Tıp Fak�ltesi, Tıbbi Mikrobiyoloji Anabilim Dalı, Adana.
5 �ukurova University Faculty of Medicine, Department of Medical Microbiology, Adana, Turkey.
* Bu �alışma, Pamukkale �niversitesi Bilimsel Araştırma Projeleri Koordinasyon Birimi tarafından (Proje no: 2017HZDP020) desteklenmiş ve �alışmanın sonu�ları 20. Uluslararası İnsan ve Hayvan Mikoloji Derneği Kongresi (20th International Society for Human and Animal Mycology-ISHAM, 30 Haziran-4 Temmuz 2018)'nde poster olarak sunulmuştur.
Makale Atıfı: Şeng�l M, Kutlu M, D�ğen A, Aksoy L, Gonca S, İlkit M ve ark. Denizli B�lgesinde Kestane (Castanea sativa) Ağacından Cryptococcus neoformans İzolasyonu. Mikrobiyol Bul 2019;53(1):61-69
�Z
Bazidiomi�et bir maya mantarı olan Cryptococcus neoformans hayvanlar ve bağışık sistemi baskılı insanlarda hayatı tehdit eden enfeksiyonlara neden olabilmektedir. �evresel ortam havasında bulunan aerosolize partik�llerin solunması ile C.neoformans/Cryptococcus gattii enfeksiyonlarının oluştuğu d�ş�n�lmektedir. Bilinen �evresel kolonizasyon odakları; kanatlı �ıkartıları, ağa�lar i�indeki ��r�yen oyuklar ve topraktır. Son yıllarda Anadolu'nun batı b�lgelerinde yapılan �evresel taramalarda Eucalyptus camaldulensis, Tamarix hispida, Platanus orientalis ve Punica granatum gibi farklı ağa�ların odunsu yapılarında C.neoformans kolonizasyonu bildirilmiştir. Eldeki verilere g�re yapılan tahminler �lkemizin batı b�lgelerinde yoğun Cryptococcus kolonizasyon odaklarının olabileceğini �ng�rmektedir. Bu �alışmada, y�ksek rakımlı b�lgelerde bulunan kestane (Castanea spp.) ağa�larında C.neoformans kolonizasyonu varlığını araştırmak ama�lanmıştır. Bu ama�la, Aydın-Denizli-�demiş b�lgesinde bulunan kestane (Castanea spp.) ağa�larında C.neoformans kolonizasyonu araştırılmıştır. �alışma, iki b�y�k nehir arasında kalan verimli, y�ksek rakımlı dağlık arazide yapılmıştır. Bu b�lge Anadolu'da yaygın kestane tarımı yapılan b�lgeler arasında yer almaktadır. Bu alanda, 2017 yılının yaz aylarında, �zerinde derin yarık veya g�vdesinde b�y�k oyuk bulunan 214 kestane ağacından �rnek alınmıştır. Ek�vyon tekniği ile alınan �rnekler bifenil ve antibiyotik i�eren Staib agar besiyerine ekim yapılmıştır. K�lt�rler on g�n s�re ile kahverengi koloni kontrol� yapılarak takip edilmiştir. Patojen Cryptococcus izolatlarını tanımlamak i�in ş�pheli koloniler konvansiyonel y�ntemler ve kanavanin glisin bromtimol agar reaksiyonu ile incelenmiştir. İzolatın polimeraz zincir reaksiyonu (PCR) tanısı i�in ITS 1-4 primerleri kullanılmıştır. Bu ama�la, STE20 (Aa), STE20 (A), STE20 (Da) ve STE20 (D) primerlerini i�eren STE20 PCR paneli ile serotip ve eşeyli şekil saptanmıştır. V8 agar besiyeri eşeyli �aprazlama k�lt�rleri i�in kullanılmıştır. �alışmada incelenen 214 ağacın yalnızca bir tanesinde (%0.47) C.neoformans izolasyonu ger�ekleşmiştir. Bu izolat ITS 1-4 dizi analizi ile doğrulanmıştır. İzolatın A serotipinde MATa şeklinde olduğu saptanmıştır. V8 agarda MATa yapısına ait bazidiyum, bazidiyosporlar ve hif yapılarındaki kelep�e "klamp" bağlantıları g�r�lm�şt�r. Bu �alışmada, Denizli'de Anadolu kestane (Castanea sativa) ağacından C.neoformans'ın ilk defa izolasyonu g�sterilmiştir. İnsan patojeni kriptokokların �evresel dağılımı ile yapılacak ileri araştırmalar, b�lgemizdeki canlılarda risk oluşturan alanların saptanmasında yardımcı olacaktır.
Anahtar kelimeler: Cryptococcus neoformans; Castanea sativa; �evresel.
ABSTRACT
Cryptococcus neoformans is a basidiomycetous encapsulated yeast that can cause life-threatening infections in immunosuppressed humans and animals. C.neoformans/Cryptococcus gattii infections are considered to be acquired via inhalation of aerosolized particles from the environment. Avian guano, decaying tree hollows and soil are known as environmental niches. In recent years, colonization of the woody structures of different trees such as Eucalyptus camaldulensis, Tamarix hispida, Platanus orientalis and Punica granatum has been reported in the environmental study of the western Anatolian region. Based on the results of previous studies, our country may have intensive Cryptococcus colonization niches in the western regions. The aim of this study was to investigate the presence of the colonization of C.neoformans niche in chestnut (Castanea spp.) trees on higher altitudes. In the study, the colonization of C.neoformans was screened on chestnut trees (Castanea spp.) in Aydın-Ödemiş-Denizli geographical area. This area consists of mountainous terrain between the fertile plain formed by two major rivers.This region is one of the widespreading areas of chestnut farming in Anatolia. Two hundred and fourteen chestnut trees that had deep fissures or trunk hollows were screened during mid-summer 2017. A swabbing technique was used, and all samples were cultured on Staib agar medium containing biphenyl and antibiotics. Cultures were checked for ten days for suspicious brown colonies. Suspicious yeast colonies were tested for the identification of pathogenic Cryptococcus by conventional methods and canavanine-glycine-bromothymol agar reactions. ITS 1-4 primers were used for strain PCR tests. We determined the mating type and serotypes by PCR analysis of the STE20 genes using STE20 (Aa), STE20 (A), STE20 (Da), and STE20 (D) primers. V8 agar medium was used for mating cultivation. Only one (0.47%) strain of C.neoformans was isolated from 214 screened trees. This strain was confirmed by ITS 1-4 sequencing. The serotype A MATa mating type was observed. Basidium, basidiospores and clamp connections in hyphal structure were noted with MATa mating on V8 agar medium. In this study, the first C.neoformans isolate from a chestnut tree (Castanea sativa) was determined from Denizli region. Further studies of distribution of human pathogenic Cryptococcus will be helpful to determine the risk areas for the living organisms in our region.
Keywords: Cryptococcus neoformans; Castanea sativa; environmental.
Geliş Tarihi (Received): 03.08.2018 - Kabul Ediliş Tarihi (Accepted): 01.11.2018
GİRİŞ
Cryptococcus neoformans ve Cryptococcus gattii �zellikle bağışık sistemi baskılanmış konaklarda hayatı tehdit eden enfeksiyonlara neden olabilen bazidiomi�et maya mantarları grubunda yer almaktadırlar. �oğunlukla odun flora ve kanatlı �ıkartılarında kolonize olmakta ve �evresel ortamlarda �oğalmaktadırlar. Kolonize oldukları �evresel ortamlarda basidiosporların ve zor şartlarda bile yaşama �zelliği bulunan maya h�crelerinin inhalasyon havasına karışması ile insanlara bulaşmaktadırlar1-4. Molek�ler araştırmalar insan kriptokokkozunun en �nemli kaynağının mayaların kolonize olduğu �evresel ortamlar olduğunu g�stermektedir5,6.
�lkemizde �evresel ortamlardaki kanatlı �ıkartılarından 1974'ten beri C.neoformans izolasyonları yapılmaktadır7-10. Ağa� florasından yapılan izolasyonlar ise 2004'ten g�n�m�ze kadar, flora farklılıklarını da i�erecek şekilde bildirilmektedir11-15. Ağa� florasından yapılan araştırmaların �oğunluğu primer odakların bulunması ve mayanın biyolojisi ile doğada yayılma �zelliklerin belirlenmesi amacıyla yapılmaktadır16. Akdeniz �lkelerinde ve Batı Anadolu sahil şeridi ile birlikte i� b�lgelerde de yaygın kolonizasyon g�r�lebileceği �ng�r�lmektedir17,18. Bu nedenle, hem doğada mayanın davranışı hem de insanlar i�in riskli b�lgelerin saptanması i�in, �eşitli b�lgelerde farklı ağa� floralarının mayanın kolonizasyonu bakımından araştırılması gerekmektedir.
Tayland'da y�ksek rakımda yetişen yerel kayıngiller ailesinden bir ağa�tan (yalancı kestane, Castanopsis argyrophylla, aile: Fagaceae) C.gattii izolasyonu bildirilmiştir19. �lkemizin de y�ksek rakımlı b�lgelerinde, kayıngiller ailesinden kestane yetiştiriciliği yaygındır ve d�nya kestane tarımında �nemli bir �retim alanıdır20. Kestane ağa�ları ile insan teması b�lgemizde tarımsal �retim nedeniyle yoğundur. Bu �alışmada, Denizli-Aydın b�lgesinde, y�ksek rakımda yetişen kestane (Castanea spp.) ağa�larında C.neoformans kolonizasyonunu araştırmak ama�lanmıştır.
GERE� ve Y�NTEM
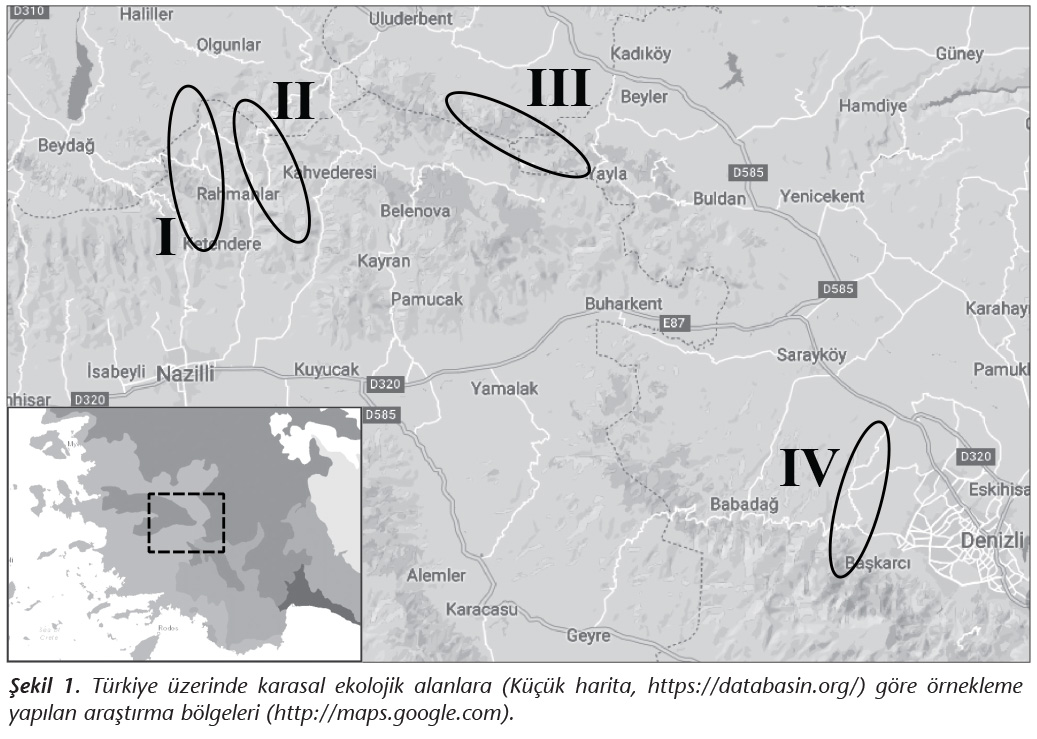
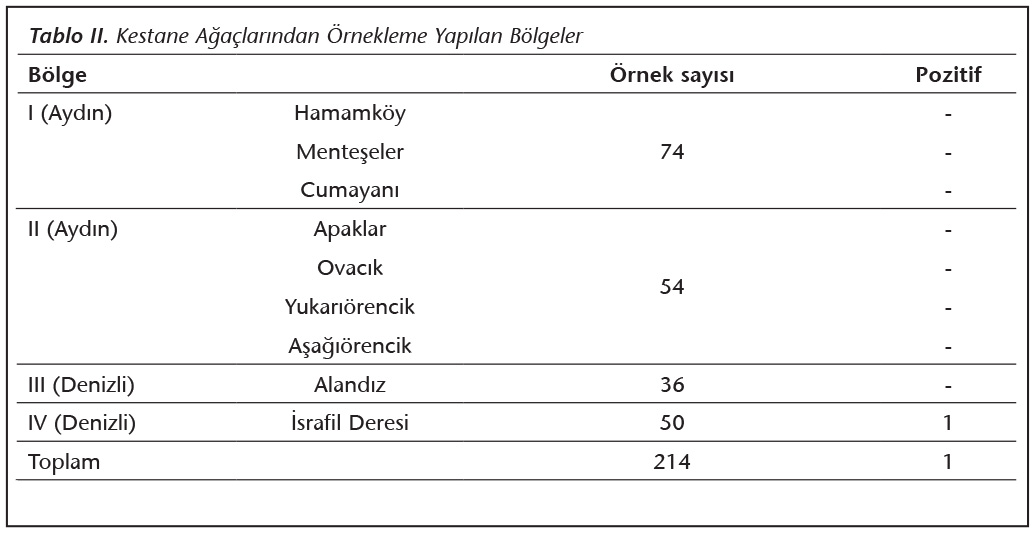
�alışma Aydın-Denizli illeri arasındaki arazinin �oğunluğu coğrafi olarak derin vadilerin dikey olarak kestiği, K���k Menderes ve B�y�k Menderes Havzaları arasında kalan b�lgede ger�ekleştirildi. Bu b�lgede, Bozdağlar �zerinde bulunan �� farklı alan ve Denizli Başkarcı b�lgesindeki bir alan, kestane (Castanea spp.) ağa�ları varlığı y�n�nden tarandı (Şekil 1) ve 224 ağa�tan Randhawa ve arkadaşları21 tarafından tanımlanan ek�vyon y�ntemi ile �rnekleme yapıldı. Tarama b�lgesinde, �zerinde derin yarık ve kovuk bulunan ağa�lar �rnekleme i�in araştırmaya dahil edildi.
�rnek Alımı ve Mikrobiyolojik Tanı
İ�erisinde taşıma ortamı (%0.4 kloramfenikol i�eren steril edilmiş %0.9 NaCl) bulunan steril ek�vyonlar �rnek alımında kullanıldı. Taşıma ortamı ile ıslatılan ek�vyon, ağa� kovuğunun ve/veya derin yarıklarının farklı b�lgelerine s�r�ld� ve �rnekler steril t�p i�ine aktarıldı. Alınan �rnekler aynı g�n i�inde laboratuvara ulaştırıldı. Ek�vyonu �ıkarılan �rnekleme t�p�, bir dakika s�reyle vortekslendi ve ��kme işlemi i�in oda sıcaklığında 30 dakika bekletildi. Sıvı y�zeyinden 100 μl alınarak %0.1 bifenil ve %0.5 kloramfenikol i�eren Staib agara ekildi. Nemli ortamda, 28�C'de 15 g�n s�reyle ink�basyona devam edildi ve g�naşırı �reme kontrol� yapıldı. Staib agarda; kahverengi pigment oluşturan kolonilerin �reaz aktivitesi, 37�C'de �reyebilme ve Dalmau agarda yapısal �zellikleri incelendi. �reaz pozitif, 37�C'de �reyebilen ve hif oluşturmayan kolonilerin kanavanin-glisin-bromtimol agar besiyerinde �remesi g�zlendi. Bu besiyerinde �reyemeyen koloniler, C.neoformans kabul edilerek molek�ler amplifikasyon ve V8 agarda eşeyli �aprazlama y�ntemleri kullanılarak ileri tanımla yapıldı. �alışmada kontrol olarak C.neoformans KN99 Aa ve C.neoformans KN99 A standart suşları kullanıldı.
Molek�ler Y�ntemlerle Tanı
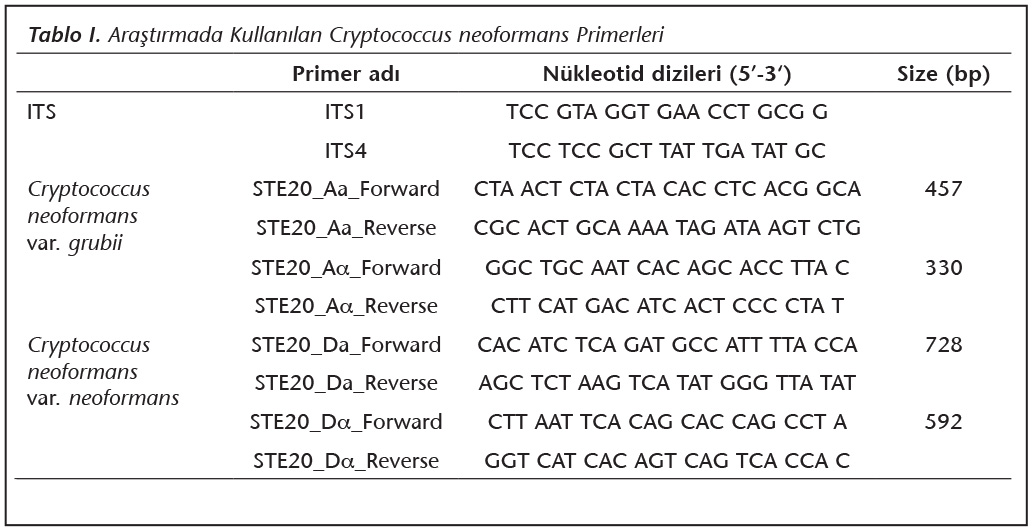
İzole edilen koloninin ve standart C.neoformans suşlarının [JEC 20 (VNIV-Da), JEC 21 (VNIV-Da), KN99 Aa ve KN99 Aa] genomik DNA ekstraksiyonu Turin ve arkadaşları22 tarafından �nerilen y�nteme g�re ger�ekleştirildi. ITS 1-4 primerleri kullanılarak PCR amplifikasyonu, toplam hacim 25 μl olacak şekilde [12.5 μl 2X karışım (Ampliqon Taq DNA Polymerase Master Mix, Odense, Danimarka); 0.5 μl her bir primerden (100 pmol/μl), 10.5 μl distile su ve 1 μl kalıp DNA] ger�ekleştirildi (Tablo I). Isı d�ng� cihazı (Techne Prime, Birleşik Krallık)'nda, PCR amplifikasyonu 95�C'te 5 dakika ilk denat�rasyonu takiben, 35 d�ng� 95�C'te 45 saniye denat�rasyon, 57�C'te 60 saniye primer birleşmesi, 72�C'te 60 saniye �oğaltma ve 72�C'de 5 dakika son uzama olarak optimize edildi. PCR amplifikasyonu sonrasında elektroforez ile %1.2'lik agaroz jelde 120 voltta 30 dakika y�r�t�len �rnekler jel g�r�nt�leme cihazında incelendi. Hedeflenen b�lgede bant g�zlenen PCR �rnekleri saflaştırıldıktan sonra, RefGen Biyoteknoloji ve Araştırma Merkezi'nde (ODT�, Ankara) dizi analizi ile doğrulandı.
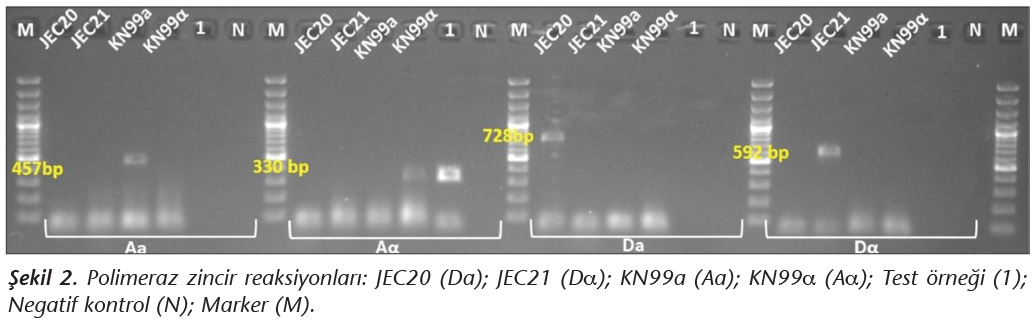
Eşeyli tipin saptanması amacıyla; JEC 20 (VNIV-Da), JEC 21 (VNIV-Da), KN99Aa, KN99Aa ve test edilen izolatın genomik DNA ekstraksiyonu, STE20 Aa, STE20 Aa, STE20 Da, STE20 Da primerleri (Tablo I) kullanılarak PCR amplifikasyonu yapıldı. STE20 Aa, STE20 A, STE20 D primerleri ile yapılan amplifikasyonda, 95�C'te 6 dakika ilk denat�rasyon sonrası, 36 d�ng� 95�C'te 45 saniye denat�rasyon, 60�C'te 45 saniye primer birleşmesi, 72�C'te 1.5 dakika �oğaltma ve takiben 72�C'de 7 dakika son uzama ger�ekleştirildi. STE20 Da primeri ile yapılan amplifikasyon 95�C'te 6 dakika ilk denat�rasyon sonrası, 30 d�ng� 95�C'te 45 saniye denat�rasyon, 50�C'te 45 saniye primer birleşmesi, 72�C'te 1.5 dakika �oğaltma ve takiben 72�C'de 7 dakika son uzama olmak �zere ısı d�ng� cihazında ger�ekleştirildi. Amplifikasyon sonrasında PCR �r�nleri elektroforez ile %1.4'l�k agaroz jelde 120 voltta 60 dakika y�r�t�lerek jel g�r�nt�leme cihazında incelendi.
Eşey �aprazlama testinde; izolat C.neoformans KN99 Aa ve C.neoformans KN99 Asuşları ile V8 agar [%5 V8 (Campbell's, Kanada), 3 mM KH2PO4, %4 agar; 1 N KOH ile pH= 5.5 ayarlandı] besiyerinde �aprazlanarak eşeyli �reme i�in karanlıkta, 25�C'te d�rt hafta s�re ile ink�basyona bırakıldı23. S�re sonunda, kontrol ve �aprazlama ekimleri, ışık mikroskobunda hif yapıları ve basidiyum/basidiyospor varlığı y�n�nden değerlendirildi.
BULGULAR
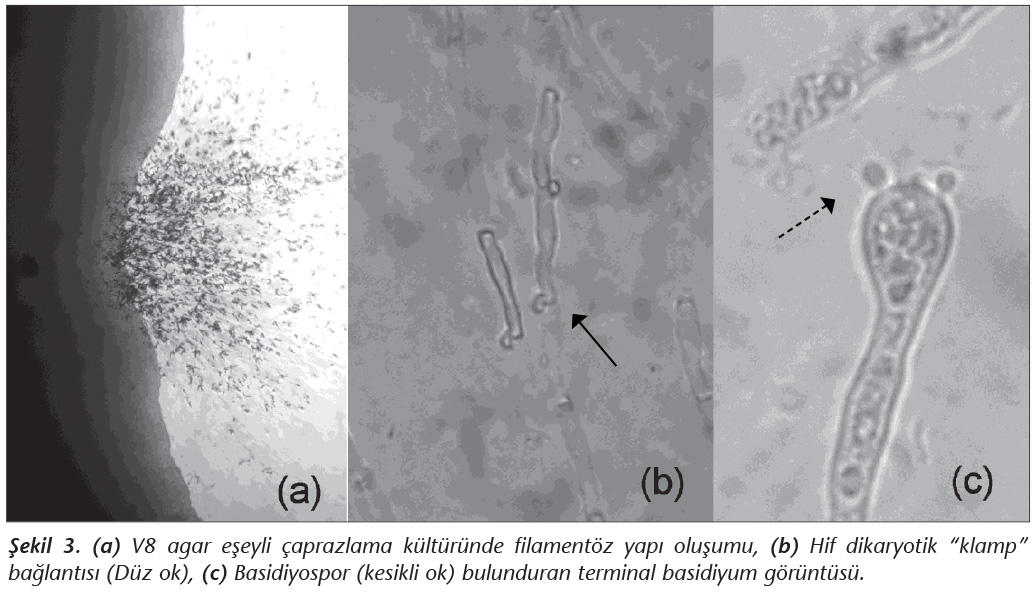
�alışmaya alınan d�rt b�lgedeki toplam 224 ağa�tan toplanan �rneklerin yalnızca birinde C.neoformans �remesi saptanmıştır. Mayanın izole edildiği b�lgenin Denizli İsrafil Deresi civarı olduğu belirlenmiştir (Şekil 1, Tablo II). İzolatın A serotipinde ve MATa genotipinde olduğu saptanmıştır (Şekil 2). V8 agarda yapılan eşeyli �aprazlama k�lt�r�nde filament�z yapı oluşumunda dikaryotik "klamp" bağlantısı ve basidiyospor bulunduran terminal basidiyum g�r�nt�s� KN99 Aa ile elde edilerek test izolatının MATa olduğu doğrulanmıştır (Şekil 3).
TARTIŞMA
Farklı �evresel ortamlarda kolonize olabilen C.neoformans'ın ekolojisine ilişkin bilgilerimiz hen�z yeterli değildir. Bu mayanın bulunduğu odunsu yapılar ile birlikte �evrenin iklimsel �zelliklerinin mayanın dağılımında �nemli rol oynadığı da kabul edilmektedir. Daha soğuk iklimlere ve yazları daha y�ksek neme uyum sağlamasından dolayı C.neoformans'ın C.gattii'ye g�re, Akdeniz �lkelerinde daha yaygın olarak bulunduğu �ne s�r�lmektedir. C.neoformans'ın dağılımı ile ilgili Cogliati ve arkadaşlarının24 Akdeniz havzasında y�r�tt�ğ� geniş katılımlı bir araştırma ve buna bağlı iklimsel t�r dağılım modellemesinde17, �lkemizin de i�inde bulunduğu riskli b�lgeler belirlenmeye �alışılmıştır. Yağışlı d�nemlerde bu mikroorganizmaların ortamda hayatlarını s�rd�rebilme yeteneklerinin farklılığı, anahtar rol oynayan bir fakt�r olarak belirtilmiştir. Benzer şekilde bağıl nemin y�ksek olduğu d�nemlerde, ortamda bulunan mikrobiyotanın değişkenliğine de bağlı olarak, C.neoformans �evresel ortamlardan daha y�ksek oranda izole edilmektedir25,26. Sunulan araştırmanın yapıldığı b�lge, K�ppen-Geiger iklim sınıflamasına g�re "CSA" (kışı ılık, yazı �ok sıcak ve kurak iklim (Akdeniz iklimi), Tsıcak ≥ 22�C) sınıfında yer almaktadır27. �lkemizde kestane tarımı genellikle 10�C �st�nde ve yıllık yağışın 1000 mm3'�n altına d�şmediği b�lgelerde yapılmaktadır20. Kestanenin daha soğuk iklimlerde yetişebiliyor olması, sunulan araştırmada da elde edilen veriye uygun olarak, C.neoformans kolonizasyonu i�in uygun bir mikroklima ortamı sağlayacağını d�ş�nd�rmektedir. Kestanenin ılıman iklim b�lgesi ağacı olması ve kurak ge�en mevsimlerin bitkiye zarar vermesinden dolayı, kestane yetişen Anadolu b�lgelerinde floranın taranması, C.neoformans'ın ekolojisinin anlaşılmasında �nemli bir �evresel yanıt olabilir. Bununla birlikte, Akdeniz havzasındaki patojen �evresel kriptokokların dağılımını inceleyen bir araştırmada 46 C.sativa ağacından �rnek alınmış, ancak �reme saptanmamıştır24. Bu durum, yapılan araştırmaların �oğunlukla iklim farklılıkları g�steren deniz seviyesi kısmında y�r�t�lmesi, yoğun kestane tarımı yapılan yerlerin ise daha i� ve ormanlık b�lgelerde bulunması nedeniyle olabilir. Cogliati ve arkadaşlarının17 Akdeniz havzasında muhtemel �evresel odakların yaygınlığını inceledikleri �alışmada, �zellikle Marmara B�lgesi i� kısımlarında, kıyı b�lgelere g�re daha y�ksek oranda C.neoformans var. neoformans g�r�lme olasılığının y�ksek olabileceği belirlenmiştir. Cogliati ve arkadaşları17 ile Acherson ve arkadaşlarının18 yaptıkları �alışmalarda C.gattii varlığının ise daha ılıman olan Ege ve G�ney Anadolu coğrafyasında olabileceği bildirilmiştir.
Araştırmada saptanan izolat, MATa eşeyli formunda olup, bu formun �lkemizde baskın olduğu �nceki �alışmalar ile de g�sterilmiştir28. D�nya �zerinde de en baskın olan form alel tipidir29. Yapılan araştırmalarda MAT lokusundaki farklılıkların saptanmasının, C.neoformans'ın ekolojisi ve vir�lansı ile bağlantısının aydınlatılmasında �nemli olduğu belirtilmektedir30. �aprazlama testlerinde de izolat KN99 Aa ile eşeyli şekil oluşturmuştur. V8 agar, Escand�n ve arkadaşlarının23 tanımladığına g�re hazırlanmış ve pH= 5.5 olarak ayarlanmıştır. Bu ortamda da bazidiyum, bazidiyospor ve kelep�e "klamp" bağlantıları g�sterilmiştir (Şekil 3). Yapılan �alışmalarda eşeyli şeklin saptanması �nemlidir. Konuyla ilgili laboratuvarlarda kullanılan V8 agar pratik, ucuz ve değerlendirmesi kolay bir besiyeri olarak tanıya yardımcı olmaktadır.
�lkemizde okaliptus (Eucalyptus camaldulensis), ılgın (Tamarix hispida), doğu �ınarı (Platanus orientalis), nar (Punica granatum) ve �am ağa�larında C.neoformans izolasyonu daha �nceki araştırmalar ile bildirilmiştir11-15. Yapılan araştırmalar ve coğrafi analizler g�z�n�ne alındığında, Marmara, Ege ve �ukurova (ve kısmen Doğu Karadeniz) b�lgelerinde insan ve hayvan patojeni bu maya mantarının araştırılması ve y�ksek kolonizasyon b�lgelerinin saptanması m�mk�nd�r17,18. Planlanarak yapılacak yeni araştırmalar hem mayanın ekolojisinin anlaşılmasına yardımcı olacak, hem de d�nyada yoğun şekilde ger�ekleştirilen �evresel kriptokok taramalarına �nemli katkı sağlayacaktır.
�IKAR �ATIŞMASI
Yazarlar bu makale ile ilgili herhangi bir �ıkar �atışması bildirmemişlerdir.
KAYNAKLAR
- Kwon-Chung KJ, Bennett JE, Wickes BL, Meyer W, Cuomo CA, Wollenburg KR, et al. The case for adopting the "species complex" nomenclature for the etiologic agents of cryptococcosis. mSphere 2017;2(1):e00357-16.
- Lin X, Heitman J. The biology of the Cryptococcus neoformans species complex. Annu Rev Microbiol 2006;60:69-105.
- Springer DJ, Saini D, Byrnes EJ, Heitman J, Frothingham R. Development of an aerosol model of Cryptococcus reveals humidity as an important factor affecting the viability of Cryptococcus during aerosolization. PLOS One 2013;8(7):e69804.
- May RC, Stone NR, Wiesner DL, Bicanic T, Nielsen K. Cryptococcus: from environmental saprophyte to global pathogen. Nat Rev Microbiol 2016;14(2):106-17.
- Chen Y, Litvintsenva AP, Frazzitta AE, Haverkamp MR, Wang L, Fang C, et al. Comparative analyses of clinical and environmental populations of Cryptococcus neoformans in Botswana. Mol Ecol 2015;24(14):3559-71.
- Kangogo M, Bader O, Boga H, Wanyoike W, Folba C, Worasilchai N, et al. Molecular types of Cryptococcus gattii/Cryptococcus neoformans species complex from clinical and environmental sources in Nairobi, Kenya. Mycoses 2015;58(11):665-70.
- Tümbay E. İzmir yöresinde Cryptococcus neoformans ve kriptokokkoz. Birinci kısım: Cryptococcus neoformans'ın doğal kaynaklarından izolasyonu. TÜBİTAK 6'ncı Bilim Kongresi, 17-21 Ekim 1977, Ankara. Tıp Araştırma Grubu Tebliğleri Tutanağı, s:839-63.
- Yıldıran ŞT, Sara�lı MA, G�nl�m A, G�n H. Isolation Cryptococcus neoformans var. Neoformans from pigeon droppings collected throughout Turkey. Med Mycol 1998;36(6):391-4.
- Karaca Derici Y, Tümbay E. İzmir ilinde doğal ve klinik Cryptococcus neoformans kökenlerinin varyete ve serotipleri. İnfek Derg 2008;22(1):53-8.
- Pelek Ş, Altınkaya S, Korkmaz UB, Ergin �. Denizli şehir merkezinde g�vercin (Columba livia) �ıkartılarında Cryptococcus neoformans varlığının araştırılması. Pam Tıp Derg 2011;4(1):21-4.
- Ergin C, Ilkit M, Hilmioğlu S, G�lbaba AG, Demirci M, Kaya S. The first isolation of Cryptococcus neoformans from Eucalyptus trees in South Aegean and Mediterranean Regions of Anatolia in Turkey despite Taurus Mountains alkalinity. Mycopathologia 2004;158(1):43-7.
- Ergin C, Kaleli I. Denizli şehir merkezinde kovuklu ağa� g�vdelerinden Cryptococcus neoformans izolasyonu. Mikrobiyol Bul 2010;44(1):79-85.
- Ergin �, Şeng�l M, Kiriş Satılmış �. T�rkiye'nin G�ney-Batı Anadolu b�lgesindeki ılgın (Genus: Tamarix L.) ağa�larında Cryptococcus neoformans kolonizasyonu takibi. Turk Mikrobiyol Cem Derg 2014;44(4):158-62.
- G�k�en H, Ergin �. Muğla-Milas il�e yerleşimi b�lgesi Eucalyptus camaldulensis ağa�larından Cryptococcus neoformans izolasyonu. Pam Tıp Derg 2014;7(2):109-12.
- Cogliati M, Zani A, Rickerts V, McCormick I, Desnos-Ollivier M, Velegraki A, et al. Multilocus sequence typing analysis reveals that Cryptococcus neoformans var. neoformans is a recombinant population. Fungal Genet Biol 2016;87:22-9.
- Randhawa HS, Kowshik T, Chowdhary A, Preeti Sinha K, Khan ZU, Sun S, et al. The expanding host tree species spectrum of Cryptococcus gattii and Cryptococcus neoformans and their isolations from surrounding soil in India. Med Mycol 2008;46(8):823-33.
- Cogliati M, Puccianti E, Montagna MT, De Donno A, Susever S, Ergin C, et al. Fundamental niche prediction of the pathogenic yeasts Cryptococcus neoformans and Cryptococcus gattii in Europe. Environ Microbiol 2017;19(10):4318-25.
- Acheson ES, Galanis E, Bartlett K, Mak S, Klinkenberg B. Searching for clues for eighteen years: deciphering the ecological determinants of Cryptococcus gattii on Vancouver Island, British Columbia. Med Mycol 2018;56(2):129-44.
- Khayhan K, Hagen F, Norkaew T, Puengchan T, Boekhout T, Sriburee P. Isolation of Cryptococcus gattii from a Castanopsis argyrophylla tree hollow (Mai-Kaw), Chiang Mai, Thailand. Mycopathologia 2017;182(3-4):365-70.
- Karadeniz V. T�rkiye'de kestane tarımı ve başlıca sorunları. Uluslararası Sosyal Araştırmalar Dergisi 2013;6(27):279-91.
- Randhawa HS, Kowshik T, Khan ZU. Efficacy of swabbing versus a conventional technique for isolation of Cryptococcus neoformans from decayed wood in tree trunk hollows. Med Mycol 2005;43(1):67-71.
- Turin L, Riva F, Galbiati G, Cainelli T. Fast, simple and highly sensitive double-rounded polymerase chain reaction assay to detect medically relevant fungi in dermatological specimens. Eur J Clin Invest 2000;30(6):511-8.
- Escand�n P, Ngamskulrungroj P, Meyer W, Casta�eda E. In vitro mating of Colombian isolates of the Cryptococcus neoformans species complex. Biomedica 2007;27(2):308-14.
- Cogliati M, D'Amicis R, Zani A, Montagna MT, Caggiano G, De Giglio O, et al. Environmental distribution of Cryptococcus neoformans and C.gattii around the Mediterranean basin. FEMS Yeast Res 2016;16(4).
- Granados DP, Casta�eda E. Isolation and characterization of Cryptococcus neoformans varieties recovered from natural sources in Bogot�, Colombia, and study of ecological conditions in the area. Microb Ecol 2005;49(2):282-90.
- V�lez N, Escand�n P. Distribution and association between environmental and clinical isolates of Cryptococcus neoformans in Bogota Colombia, 2012-2015. Mem Inst Oswaldo Cruz 2016;111(10):642-8.
- �zt�rk MZ, �etinkaya G, Aydın S. K�ppen-Geiger iklim sınıflandırmasına g�re T�rkiye'nin iklim tipleri. Cografya Dergisi 2017;35:17-27.
- Sara�li MA, Yıldıran ŞT, Şener K, Gonlum A, Doganci L, Keller SM, et al. Genotyping of Turkish environmental Cryptococcus neoformans var. neoformans isolates by pulsed field gel electrophoresis and mating type. Mycoses 2006;49(2):124-9.
- Kwon-Chung KJ, Bennett JE. Distribution of alpha and alpha mating types of Cryptococcus neoformans among natural and clinical isolates. Am J Epidemiol 1978;108(4):337-40.
- Fu C, Sun S, Billmyre RB, Roach KC, Heitman J. Unisexual versus bisexual mating in Cryptococcus neoformans: Consequences and biological impacts. Fungal Genet Biol 2015;78:65-75.
İletişim (Correspondence):
Prof. Dr. �ağrı Ergin,
Pamukkale �niversitesi Tıp Fak�ltesi,
Tıbbi Mikrobiyoloji Anabilim Dalı,
Denizli, T�rkiye.
Tel (Phone): +90 258 296 2491,
E-posta (E-mail): cagri@pau.edu.tr