Hepatit C Virusunun Genomik Varyasyonları ve Kliniğe Etkileri
Clinical Impact of Hepatitis C Virus Genomic Variations
Koray ERGÜNAY1, Hakan ABACIOĞLU2
1 Hacettepe Üniversitesi Tıp Fakültesi, Tıbbi Mikrobiyoloji Anabilim Dalı, Ankara.
1 Hacettepe University Faculty of Medicine, Department of Medical Microbiology, Ankara, Turkey.
2 İzmir Ekonomi Üniversitesi Tıp Fakültesi, Tıbbi Mikrobiyoloji Anabilim Dalı, İzmir.
2 Izmir University of Economics Faculty of Medicine, Department of Medical Microbiology, Izmir, Turkey.
* Koray Ergünay, Alexander Von Humboldt Vakfı “Georg Forster Research Fellowship (HERMES)” programı tarafından desteklenmektedir (2015).
ÖZ
Hepatit C virusu (HCV), tüm dünyada yaygın olarak bulunan, 110 milyonu aşkın kişiyi enfekte etmiş, ciddi bir halk sağlığı tehdidi oluşturan kronik hepatit etkenidir. Maruziyetin ardından, kronik hastalık riski yüksek olan ve koruyucu bir aşısı bulunmayan HCV; siroz, transplantasyon gerektiren karaciğer yetmezliği ve hepatoselüler karsinomun önde gelen enfeksiyöz etkenleri arasında yer almaktadır. Replikasyon mekanizmaları ve konaktaki enfeksiyon dinamikleri nedeniyle, virus, enfekte kişilerde türümsüler (quasispecies) olarak tanımlanan, birbirine çok benzer genetik varyantları içeren bir popülasyon halinde bulunur. HCV enfeksiyonlarında patogenez ve hastalığın ilerleyişi, virusa ait genetik çeşitlilik ve türümsü kompozisyonlarında izlenen değişikliklerle doğrudan ilgilidir. Virus, yüksek düzey çeşitlilik gösteren varyant havuzundan, iç ya da dış kaynaklı seçici baskıların varlığında replikasyonunu sürdürebilecek mutantların oluşumu ile, hastalık patogenezini devam ettirir. Enfeksiyon süresince, türümsüler birçok değişikliğe uğrar ve konağın hümoral ve hücresel yanıtından kaçan mutantlar, antiviral tedaviye karşı da dirençli mutantların oluşumu için zemin hazırlar. Yeni jenerasyon dizileme tekniklerinin de katkısıyla, HCV'nin kökeni ve epidemiyolojisi, ayrıca hastalık süreçlerinde izlenen genomik çeşitlilik üzerine birçok güncel bilgi elde edilmiştir. Bu derlemede, HCV'nin genetik çeşitliliğini oluşturan temel mekanizmalar ve klinik gidişle bağlantılı olarak konakla olan etkileşimler; virus ve konak yönünden hümoral ve hücresel immün yanıt, enfeksiyonun erken ve geç dönemlerindeki değişiklikler ve kronik enfeksiyona ilerleme süreci açılarından ele alınmaktadır. Ek olarak virus epidemiyolojisi ve moleküler evrimi üzerine güncel bulgular, türler arası ve popülasyon perspektiflerinden yorumlanmaktadır. Doğrudan etkili antivirallere dayalı rejimlerin uygulamaya girdiği günümüzde, genetik varyasyonlar ve tedaviye direnç ile ilişkileri, güncel saptama yöntemleri de özetlenerek tartışılmaktadır.
Anahtar sözcükler: Hepatit C virusu; HCV; türümsü; çeşitlilik; varyasyon.
ABSTRACT
Hepatitis C virus (HCV) is a globally-dispersed agent of chronic hepatitis with a significant public health threat, affecting over 110 million individuals throughout the world. The increased risk for chronicity after exposure and the lack of a protective vaccine make HCV is a leading infectious cause of cirrhosis, liver failure requiring transplantation and hepatocellular carcinoma. The replicative process and infection dynamics in the host enable HCV to generate an array of closely-related but non-identical genetic variants known as quasispecies in the infected individuals. Pathogenesis and outcome in HCV infections are directly affected by the virus genetic heterogeneity, reflected as the emergence of quasispecies in infected individuals. The evolution of these highly-diverse viral populations in the host directly influences the disease course, via providing a pool of variants capable of resuming viral replication under extrinsic and/or intrinsic selective pressures. Viral quasispecies go through several alterations during the course of the infection, and provide a background for the selection of escape mutants from the host humoral and cell-mediated immune responses and antiviral treatment. Supported by the robust next generation sequencing techniques, recent studies have provided significant insights on the genomic diversity and progression as well as on the origin and the epidemiology of HCV. This review provides an overview of the mechanisms of HCV genetic variability, and the interactions with the host, that affects clinical disease, covering viral and host determinants of humoral and cell-mediated immune responses, alterations during the early and late stages of the infection and disease progression leading to chronicity. In addition, current findings in virus evolution and epidemiology were briefly interpreted from the inter-species and population perspectives. The impact of viral genomic heterogeneity on antiviral treatment in the era of direct-acting agents is also discussed, along with an overview of current methods employed for the characterization of viral diversity.
Keywords: Hepatitis C virus; HCV; quasispecies; diversity; variation.
Geliş Tarihi (Received): 11.08.2015 • Kabul Ediliş Tarihi (Accepted): 16.09.2015
GİRİŞ
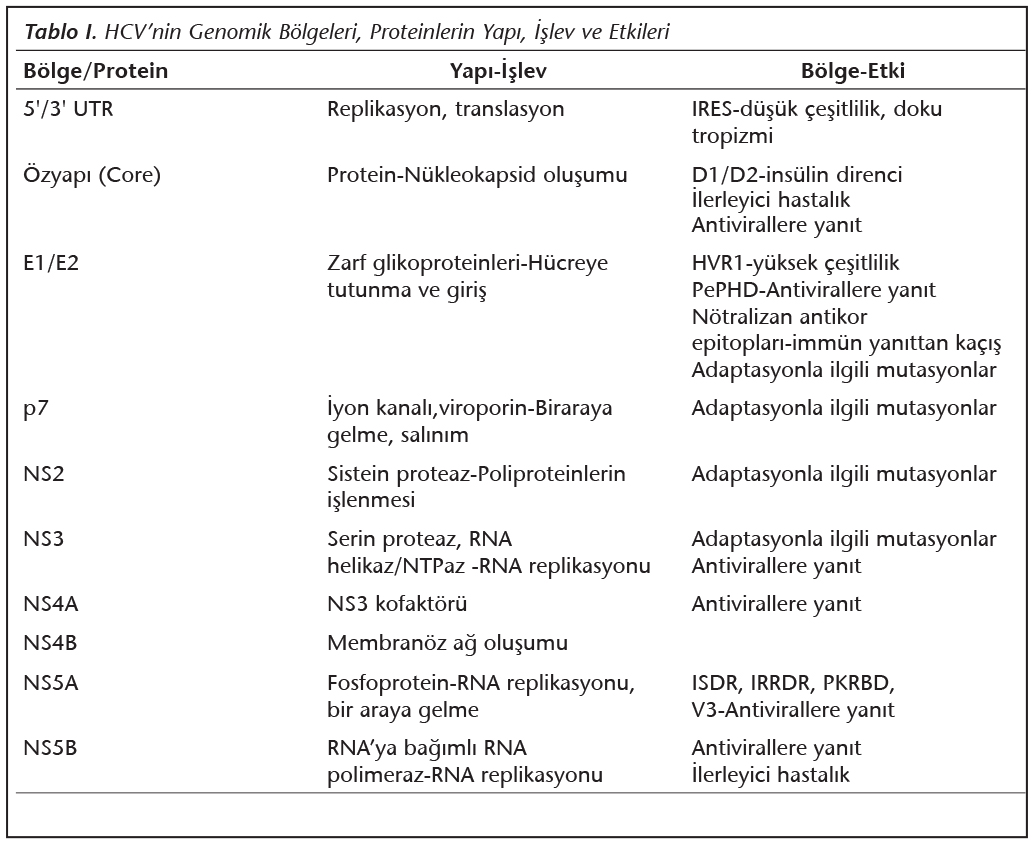
Hepatit C virusu (HCV) tarafından oluşturulan hepatitler, tüm dünyayı etkileyen ciddi bir halk sağlığı sorunudur1. Flaviviridae ailesi Hepacivirus cinsinde sınıflandırılan zarflı bir virus olan HCV, tek iplikli, pozitif polariteli yaklaşık 9.6 kilobaz içeren bir RNA genomuna sahiptir2 (Şekil 1). Viral genom, 5' ve 3' uçlarda translasyona uğramayan bölgelerle (untranslated regions; UTR) sınırlandırılmış, tek bir açık okuma çerçevesi (open reading frame; ORF) içerir. Viral poliprotein, hücresel ve viral proteazlar tarafından kesilerek olgun proteinler oluşturulur. Virusa ait yapısal proteinler; özyapı (core; C) proteini ile zarf glikoproteinleri olan E1 ve E2'dir. Konak hücrede viral replikasyonda rol oynayan yapısal olmayan proteinler ise; proteaz (NS2, NS3 ve NS4A), helikaz (NS3) ve RNA'ya bağımlı RNA polimeraz (NS5B) gibi çeşitli işlevlere sahiptir2 (Tablo I).
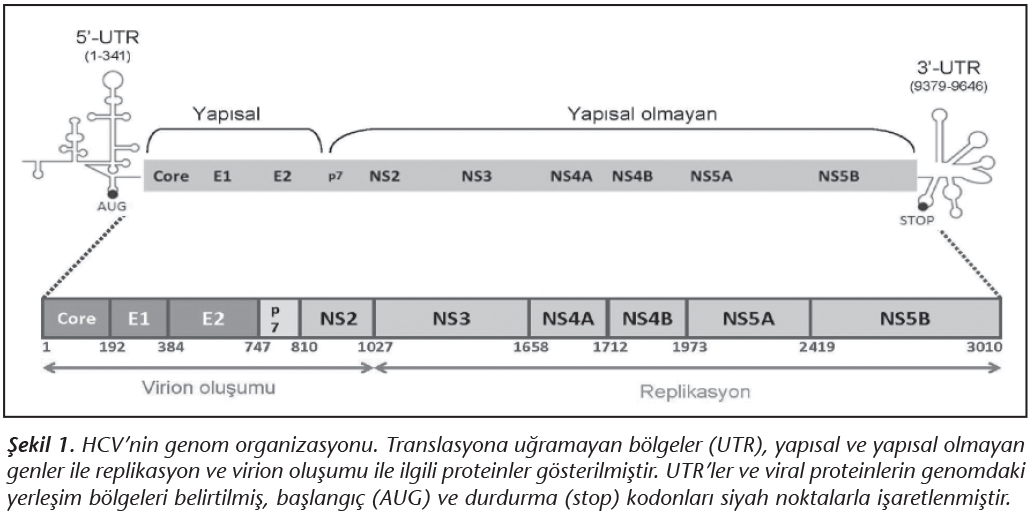
HCV'nin ayırt edici özelliklerinden birisi, konak organizmada sıradışı bir genetik çeşitlilik oluşturabilme yeteneğidir3. Bu özellik, enfeksiyonun immün yanıt varlığında dahi kronikleşmesi ve antiviral ilaç direnci gibi önemli sonuçlar doğurmaktadır. Son yıllarda yeni jenerasyon dizi analizi yöntemlerinin de yaygınlaşmasıyla, HCV enfeksiyonlarında ortaya çıkan genetik çeşitlilik ve sonuçları, ayrıntılı olarak incelenebilmiştir4. Bu derlemede, farklı perspektiflerden HCV'nin genetik çeşitliliği, sorumlu mekanizmalar ile hastalığın ilerleyişi, klinik gidiş ve tedavi ile doğrudan ilişkili olan değişimler, güncel bilgiler ışığında ele alınmaktadır.
HCV'DE GENETİK ÇEŞİTLİLİK MEKANİZMALARI
HCV enfeksiyonlarında tek konakta (intrahost) ya da farklı konaklarda (interhost) izlenen genetik çeşitliliğinin en önemli kaynağını, viral replikaz enzimine bağlı mutasyonlar oluşturur3. HCV replikazı NS5B geninden kodlanır ve RNA'ya bağımlı RNA polimeraz (RbRp) aktivitesi gösterir. Diğer RNA polimerazlar gibi HCV replikazı da, yeni sentezlenen zincirde ortaya çıkabilecek hataları düzeltebilecek 3'-5' ekzonükleaz aktivitesinden yoksundur ve yüksek hata oranına sahiptir. HCV için mutasyon oranı nükleotid başına 104-105 değişiklik olarak hesaplanmakta; bu da her replikasyon döngüsünde 1-10 genomda mutasyon olacağı anlamına gelmektedir5,6. Ayrıca, HCV'de izlenen kısa replikasyon döngüsü, enfekte kişilerde yarı ömrü 3-5 saat olan 1012 yeni virion ve beraberinde birçok mutantın oluşmasına olanak sağlamaktadır. Konaktaki virus popülasyonunun büyüklüğü de, varyasyon oluşumunu doğrudan etkilemektedir. Bu nedenle, immün yanıt ya da ilaç tedavisi sonucu replikasyonun önemli düzeyde baskılanmış olması, varyantların ortaya çıkma ihtimalini düşürmektedir3.
Yüksek mutant oluşturma kapasitesine rağmen, ortaya çıkan mutantlar konakta birçok adaptasyon ve seçilim süreçlerinden geçmek zorundadır6. İşlevsel olarak genomik çeşitliliği sınırlandıran en önemli faktör, replikasyonun başarılı bir şekilde gerçekleşebilmesi için korunması gereken dizilerin varlığıdır. HCV genomunda iyi korunmuş bölgeler, 5' ve 3' “untranslated” bölgeler ve 5'UTR'de yer alan ribozom tanıma bölgesi (internal ribosome entry site; IRES)'dir7. Buna karşın, birçok önemli antijenik epitop ve çok değişken bölge (hypervariable region; HVR) içeren E1/E2 bölgeleri, en yüksek çeşitlilik ve seçilim hızına sahiptir. E2 bölgesinde 27 aminoasitlik HVR1 bölgesi, tüm HCV genomundaki en yüksek varyasyon izlenen bölgedir. Enfekte kişilerde zaman içerisinde oluşan değişiklikler de belirgin olarak HVR1'de yoğunlaşmaktadır. Bunun dışında, immün yanıt ve antiviral ilaçların seçici baskısı da HCV genetik çeşitliliği ile doğrudan ilişkilidir3,8,9.
Ortaya çıkan mutasyonların, viral replikasyon üzerindeki olumsuz etkilerini azaltmak ya da ortadan kaldırmak için dengeleyici (kompansatuar) mutasyonlar ve aminoasit değişiklikleri ortaya çıkar. Aminoasit ko-varyasyonu kavramı içinde değerlendirilen bu değişiklikler, insan immün yetmezlik virusu (HIV)-1 ve hepatit B viruslarında da tanımlanmıştır10. HCV'de, NS3 ve NS4A mutasyonlarına bağlı NS5A fosforilasyonun modülasyonu ile replikasyonun artışı; p7 ve NS2 mutasyonları ile biraraya gelme defekti olan kor proteini mutantlarının kurtarılması; kimerik viruslarda E1, p7, NS2 ve NS3 bölgelerindeki koordineli mutasyonların, enfekte hücrede virus oluşumu ve salınımını artırması gibi değişiklikler izlenir. İyi tanımlanmış HCV ko-varyasyon ağları sayesinde, antiviral tedavi alan olgularda ilaç direnci gelişiminin öngörülmesi mümkündür10,11.
Diğer RNA viruslarında olduğu gibi, genetik çeşitliliğe katkıda bulunabilecek bir diğer mekanizma rekombinasyondur. Enfekte kişilerde çeşitli rekombinant izolatlar tanımlanmış; rekombinasyonun ortaya çıkması için uygun ortam hazırlayan karışık enfeksiyon ve süperenfeksiyonlar saptanmıştır. Bununla birlikte güncel veriler, rekombinasyonun, tüm virus genotipleri için geçerli ve yaygın izlenen bir mekanizma olmayabileceğini işaret etmektedir13.
TÜRLER ARASI DÜZEYDE HCV ÇEŞİTLİLİĞİ: VİRUSUN KÖKENLERİ
HCV ve GB virus B, uzun süre Flaviviridae ailesi Hepacivirus cinsinin yegane üyeleri olmuşlardır. Buna karşın; köpekler, atlar, kemirgenler, yarasalar ve büyükbaş hayvanlarda saptanan yeni viruslar, bu cinsin üyelerinin, birçok türde enfeksiyon oluşturabileceğini ve türler arası muhtemel geçiş olaylarının varlığını ortaya koymuştur14,15. HCV'nin kaynağı, çeşitli nedenlerle insan-dışı primatlar olarak kabul edilmiş; ancak farklı maymun türlerinde yapılan taramalar, yakın dönemde Colobus guereza maymunlarında saptanan yeni bir hepasivirus dışında, bu görüşü desteklememiştir. Buna karşın, Guereza hepasivirusu, GB virus B ile Afrika ve Amerika'da kemirici ve yarasalarda saptanan hepasiviruslara, HCV'ye göre daha yüksek genetik benzerlik göstermektedir. HCV'nin en yakın akrabası olarak, atlarda saptanan ve benzer viral kinetiklere sahip akut/kronik hepatit etkeni olan “Non-Primate Hepacivirus” (NPHV) izlenmektedir. Genomik olarak HCV'ye yaklaşık %50 homoloji gösteren NPHV'de, özellikle NS3 and NS5B bölgelerinde yüksek düzeyde benzerlik saptanmıştır. Bu bulgular üzerine, immunoblot testlerinde sınırda (intermediate) sonuç alınan kan donörlerinde NPHV dizileri araştırılmış, ancak olumlu sonuç elde edilememiştir. Bununla birlikte, NPHV proteazının, insanda doğal bağışıklıkta rol alan MAVS ve TRIF moleküllerini parçalayabilme özelliği, zoonotik bulaş ihtimalini destekler niteliktedir. Eldeki veriler, henüz HCV'nin atası olarak kabul edilebilecek bir hepasivirusa işaret etmemektedir14,15.
POPÜLASYON DÜZEYİNDE HCV ÇEŞİTLİLİĞİ: TİPLER, ALT TİPLER ve YAYILIM
Güncel kriterlere göre HCV izolatları, nükleotid düzeyinde %30-35 farklılık gösteren yedi genotip halinde gruplandırılmakta; bunlardan genotip 1 ve 3, tüm dünyada izlenen HCV enfeksiyonlarının önemli bir kısmını oluşturmaktadır16. Ek olarak, en az %15 genomik varyasyon gösteren 67 doğrulanmış ve 20 muhtemel alt tip bulunmakta; genotip 5 ve 7 tek bir alt tipe sahipken, diğer genotiplerde en az yedi alt tip izlenmektedir. Çeşitli genotip/alt tipler arasında rekombinasyonlar da tanımlanmış olup, genotip 2k/1b popülasyonları ve olgu bazlı genotip 2/5, 2b/1b, 2b/1a, ve 2i/6p rekombinantları gösterilmiştir13. Genotip/alt tipler, coğrafi ve çeşitli epidemiyolojik özelliklere göre çeşitlilik göstermekle birlikte, persistans ve hastalık ilerleyişi açısından önemli fark göstermemektedir.
Filogenetik analiz ve modellemeler, HCV'nin küresel yayılımının, 20. yüzyılda, muhtemelen AIDS epidemisinden önce meydana geldiğine işaret etmektedir14. Virusun başlıca parenteral yolla bulaşması, sterilize edilmemiş iğneler ve yaygın aşılama kampanyaları, tarama öncesi transfüzyonlar ve damar içi ilaç kullanımının, yayılımda etkili olduğunu düşündürmektedir. Farklı bölgeler ve risk faktörlerine göre genotip/alt tip analizleri de bu görüşü destekler niteliktedir. Arşivlerde 1970 yılı öncesine ait örneklerin azlığı, virus filogenetiği ve çeşitliliği üzerindeki çalışmaları olumsuz etkilemektedir14.
BİREYSEL DÜZEYDE HCV ÇEŞİTLİLİĞİ: TÜRÜMSÜLER
Enfekte kişilerde HCV varyasyonları; hataya açık replikaz enzimi ve hızlı replikasyon döngülerine bağlı olarak, aynı olmayan ancak birbirine benzerlik gösteren bir genomlar topluluğu, yani türümsüler, olarak izlenir3,8. Türümsüler, konağın seçici baskısına karşı dinamik bir süreçte değişim gösteren bir mutant spektrumu oluşturur. Bunlar aynı zamanda, virus replikasyonunu kısıtlayan immün yanıt ve antiviral tedavi gibi çevresel etkilere karşı da hızlı bir adaptasyon sağlar. Enfekte kişiler ve dokularda, HCV'nin türümsü kompozisyonu ve mutant spektrumu farklılıklar gösterir8.
HÜMORAL İMMÜN YANIT ve HCV VARYASYONLARI
Hastalarda, hayvanlarda ve in vitro modellerde, HCV'nin birçok epitopuna karşı nötralizan antikor yanıtı saptanmaktadır17. Birçok çalışma, HVR1 bölgesinin, viral nötralizasyonda kritik öneme sahip bazı doğrusal epitopları içerdiğini ortaya koymuştur17,18. Hastalığın akut döneminde ortaya çıkan HVR1 bölgesine karşı türe özgül antikorlar, viral replikasyonun kontrolü ve enfeksiyonun sınırlandırılması ile ilişkilidir. HVR1'in, daha korunmuş epitopların tanınmasını önleyerek, enfeksiyonun erken dönemde temizlenmesine engel olduğu bilinmektedir19. Virus nötralizasyonu için önemli bir hedef olması nedeniyle HVR1, seçici immünolojik baskı altındadır ve HVR1 varyantlarının oluşumu ile antikor kaçış mutantları ortaya çıkmaktadır. Enfekte kişilerden alınan takip serumları, HCV'ye karşı oluşan antikorların, dolaşımda bulunan virusları eş zamanlı olarak nötralize edemediğini, ancak önceki virus suşlarına özgül olduğunu göstermektedir20,21. HVR1'e ek olarak, E2 bölgesinde de, varyasyonların oluşumu ile nötralizasyondan kaçabilen konformasyonel epitoplar bulunmaktadır. HCV'ye karşı antikor yanıtının anlaşılması, enfeksiyonun kontrolü ve aşı geliştirilmesi açısından önemlidir. Eldeki veriler, türümsü çeşitliliğine dayanan immünolojik kaçışın, bu yolda önemli bir engel teşkil ettiğini vurgulamaktadır18.
HÜCRESEL İMMÜN YANIT ve HCV VARYASYONLARI
İnsan ve şempanzelerde, kendini sınırlayan HCV enfeksiyonlarında, sürekli ve güçlü bir CD4 ve CD8 T hücre yanıtı saptanmaktadır17,22. Bununla birlikte, virusa özgül T hücre yanıtı, her zaman virus replikasyonunun kontrolünde başarılı olamamakta; enfeksiyonun prognozu, kişiden kişiye çeşitli immünolojik mekanizmalarla ilişkili olarak farklılık göstermektedir. HCV'nin, T hücre yanıtını etkisiz kılmasındaki önemli bir mekanizma, kaçış mutantlarının ortaya çıkmasıdır17,23. Viral antijenik peptidlerin işlenmesi, MHC ile bağlanması ve CD4/CD8 T hücreleri tarafından tanınmasını, olumsuz yönde etkileyen mutantlar tanımlanmış ve ayrıca, bilinen CD8 T hücre epitoplarının önemli bir kısmında mutasyonların oluşabileceği gösterilmiştir23. Şempanze modelinden elde edilen veriler, T hücre yanıtından kaçışta, birden fazla epitopta değişikliğin olması gerektiğini düşündürmektedir22. MHC sınıf I tarafından tanınan HCV epitoplarında, CD8 T hücre yanıtında bozulmaya ve hastalığın ilerlemesine yol açan çeşitli mutasyonlar tanımlanmıştır. Buna ek olarak, T hücrelerinin seçici baskısı en yoğun olarak enfeksiyonun erken safhalarında izlenmesine rağmen; kronik HCV olgularında da, bulaşmadan yıllar sonra kaçış mutantlarının ortaya çıkabildiği bildirilmiştir. CD4 T hücre epitoplarında da mutasyonlar izlenebilmektedir. NS3 bölgesinde yer alan bir Th1 epitopunda meydana gelen belirli aminoasit değişikliği, CD4 proliferasyonunu etkilemekte ve sitokin salınım paternini Th1'den Th2 yönüne değiştirebilmektedir. Şempanze modelinde de, CD4 T hücre epitoplarının tanınmasını bozan NS3 ve NS5A mutasyonları tanımlanmıştır17,22,23.
KONAĞA ADAPTASYON ve İLERLEYİCİ HASTALIKTA HCV VARYASYONLARI
HCV enfeksiyonlarında gidiş ve sonucu belirleyen önemli faktörlerden birisi, virusun adaptasyon yeteneği ile konak immün yanıtının etkileşimidir ve bu etkileşim türümsü oluşumu ile doğrudan ilgilidir4,17. Türümsü kompozisyonundaki değişiklikler, bulaşma sonrası yeni konağa ilk adaptasyon ve karaciğerde izlenen histolojik değişiklikler ile de ilişkili bulunmuştur24,25. Perinatal olarak enfekte yenidoğanlarda, ilk 3 ayda izlenen monoklonal HCV popülasyonu, 6. ayda birçok mutant genom içeren yeni popülasyona dönüşmekte; bu değişim hümoral immün yanıtın devreye girişi işe eşzamanlı olarak ortaya çıkmaktadır26. Ardından, genetik çeşitlilikte yeniden değişim olmakta ve 19. ayda konsensus genomlar farklılaşmış olarak izlenmektedir. Transfüzyon ile oluşan bulaşmalarda, HCV türümsülerinin erken dönemdeki değişiklikleri, erişkin olgularda akut enfeksiyonun prognozu ile ilişkili bulunmuştur. Olgularda genetik çeşitlilikte belirgin azalmalar, enfeksiyonun kontrolü ve akut hepatitin iyileşmesi ile korelasyon göstermekte ve ortaya çıkan varyasyonların hemen tamamının HVR1 bölgesinde, antikor serokonversiyonu ile eşzamanlı olduğu dikkati çekmektedir. Yapılan bir çalışmada, akut HCV hepatitinde genetik çeşitliliği sınırlandıran iki dönem tespit edilmiştir24. Bunlardan ilki, bulaşma ile ilgilidir ve vericinin türümsü repertuarındaki bazı varyantların başarılı şekilde yeni konağa adaptasyonu ile ortaya çıkar. İkinci kısıtlama ise, maruziyetin yaklaşık 100. gününde özgül antikorların saptanabilir düzeye gelmesiyle oluşur; bu dönemde genetik çeşitlilikteki azalmayı, yeni varyantların oluşumu ve kronikleşme takip eder. Virusun temizlenmesinden önce ortaya çıkan genetik çeşitlilikte azalma, interferon tedavisine olumlu yanıt veren kronik enfeksiyonlarda da gösterilmiştir. Bu bulgular, klinik safhadan bağımsız olarak kontrol altına alınan HCV enfeksiyonlarında, virusa ait genetik çeşitlilikte azalma olduğunu göstermektedir. Yeni oluşan varyasyonlar ise sıklıkla HVR1 bölgesinde izlenmekte ve immün yanıta bağlı seçilimin etkisini vurgulamaktadır3,4.
Kronik HCV olgularındaki incelemeler; ilerleme izlenen olgularda, yetersiz immün yanıt ve/veya virülansı yüksek genomların baskın hale gelmesi sonucu, mutant spektrumunda bir daralmaya işaret etmektedir3,4,27. Bu durum, en belirgin olarak özyapı ve HVR1 bölgelerinde ortaya çıkmaktadır. Histolojik ilerleme ve fibrozis izlenen olgulardaki çeşitli analizler, HCV türümsülerinde homojenleşme ve HVR1 dizilerinde azalmış çeşitlilik durumunu işaret etmektedir8,25. Benzer durum fulminan hepatit gelişen olgularda da izlenmekte; yüksek düzey viremiye rağmen kısıtlı HVR1 çeşitliliği ortaya çıkmaktadır. HCV'nin konağa adaptasyonu ve seçilimi ile ilgili diğer veriler, son dönem kronik hepatit ve transplantasyon olgularından elde edilmiştir8,27.
Transplantasyon işlemi de, türümsüler üzerine seçici bir etki yapar ve varolan varyantların az bir kısmı yeni karaciğerde enfeksiyon oluşturabilir. Buna bağlı olarak, işlem sonrasında birinci aya kadar viral popülasyonlarda kısıtlı bir varyasyon izlenir. Tekrarlayan hastalığın şiddeti, transplantasyon öncesi saptanan varyantların persistansı ve azalmış genetik çeşitlilik ile korelasyon gösterir. Transplantasyon sonrası varyasyonda artış, sıklıkla daha hafif hastalığa işaret eder8,28.
HCV genomundaki bazı varyasyonların belirli hasta gruplarında daha sık izlendiği ve patogenezle ilişkili olabileceği rapor edilmektedir8,25. Genotip 3 ile enfekte kişilerde hepatik steatoz, genotip 1'e göre daha yaygındır. Özyapı proteininin D1 bölgesinde 70. aminoasidi etkileyen polimorfizmler, genotipten bağımsız olarak insülin direnci, steatoz ve hepatoselüler karsinom ile ilişkilidir. D2 bölgesindeki belirli değişikliklerin de, steatoz ve neoplastik progresyonu etkilediği düşünülmektedir25. Türümsüler üzerinde yapılan çalışmalar da benzer bulguları ortaya koymaktadır3,8.
BÖLÜMLEŞME ve HCV VARYASYONLARI
HCV'nin temel hedefi karaciğer olmasına karşın, kan hücreleri, lenfoid doku ve merkezi sinir sisteminde de enfeksiyon oluşturduğunu gösteren kanıtlar bulunmaktadır8. İmmün sistemi baskılanmış kişiler ya da tekrarlayan maruziyetlerde karaciğer dışı replikasyon daha belirgindir ve bu bölgeler transplantasyon sonrası reenfeksiyonlara da kaynak oluşturmaktadır. Çalışmalar, enfekte kişilerin farklı dokularında farklı HCV genotipleri ve türümsü kompozisyonlarının bulunduğunu ortaya koymuştur8. HIV ve HCV ile koenfekte kişilerde, plazma ve karaciğerde HVR1 çeşitliliği farklılık göstermektedir. IRES dizi analizlerinde, karaciğer harici dokulara adaptasyonla ilişkili olabilecek mutasyonlar saptanmıştır. Hepatoselüler karsinom örneklerinde de, tümör dokusunda bulunan türümsüler, çevre dokulara göre daha yüksek genetik çeşitlilik göstermektedir3,8.
ANTİVİRALLER ve HCV VARYASYONLARI
Kronik HCV'de standart tedavi yaklaşımı olarak, pegile interferon (IFN)-alfa ve ribavirin kombinasyonu yaygın olarak kullanılmaktadır; ancak tedaviye yanıt oranları istenilen düzeyde değildir29. Son yıllarda geliştirilen “doğrudan etkili antiviral (DEA)”ler, HCV tedavi yaklaşımları ve kalıcı virolojik yanıt oranlarına önemli katkılar yapmıştır. Güncel DEA grupları arasında; viral proteaz (NS3-4A), replikaz aksesuar proteini (NS5A) ve replikaz (NS5B) inhibitörleri bulunmaktadır. Standart ilaçlar ile birlikte DEA ya da IFN içermeyen DEA kombinasyonları, çeşitli tedavi rejimleri olarak denenmektedir29.
IFN + ribavirin tedavisini etkileyen viral faktörler arasında, enfeksiyonu oluşturan genotipin de önemi vardır. Genel olarak genotip 1a, 1b ve 4; 2 ve 3'e göre tedaviye daha dirençlidir. Kalıcı virolojik yanıt, en düşük olarak genotip 1 izolatlarında izlenirken; genotip 4'de orta düzey; 2, 3, 5 ve 6'da daha yüksek düzeyde izlenmektedir. IFN-temelli tedavi yanıtı ile ilişkili varyasyonlar, sıklıkla özyapı, E2 ve NS5a bölgelerinden rapor edilmektedir. Özyapı bölgesinde R70Q ve/veya L91M değişimleri, genotip 1b ile enfekte olgularda, zayıf virolojik yanıt göstergesi olarak belirlenmiştir. “PKR/eIF-2a phosphorylation homology domain” (PePHD) bölgesini oluşturan 12 aminoasitlik dizi ve çevresindeki bazı bölgelerde izlenen mutasyonların, IFN yanıtı ile ilişkisi tüm çalışmalarda doğrulanamamıştır. HCV genotip 1 izolatlarında belirli “PKR binding domain” (PKRBD) mutasyonu taşıyan olgularda, IFN tedavisine uzun dönemli olumlu yanıt rapor edilmiştir. Genotip 1b viruslarında, NS5a'nın karboksil ucuna yakın olarak konumlanan “IFN/RBV resistance-determining region” (IRRDR) bölgesindeki genetik heterojenite, IFN + ribavirin kombinasyonu uygulanan olgularda, kalıcı virolojik yanıtın öngörülmesi için bağımsız bir faktör olarak bildirilmiştir. NS5a ISDR (interferon sensitivity determining region) bölgesindeki mutasyonlar ve tedavi yanıtı ise, viral genotip, coğrafi bölge ve mutasyon sıklığına ek olarak, özyapı gibi genomdaki diğer bölgelerdeki değişikliklerle ilişkili görünmektedir. NS5a'da yer alan V3 bölgesindeki değişiklikler de, bazı gruplarda tedavi yanıtı ile bağlantılı olarak izlenmiş; bunun muhtemel nedeninin, NS5a'nın PKRBD/ISDR gibi fonksiyonel bölgelerinde ortaya çıkan modifikasyonlar olduğu öne sürülmüştür. V3 bölgesinde izlenen mutasyonların, konağın cinsiyeti ile ilişkisi ve kadınlarda daha az olması, konak ve virusun genetik özelliklerinin etkileşimini gösteren ilginç bir bulgudur4,8,11,30,31.
DEA tedavisi sırasında seçilime uğrayan, farklı ilaç sınıflarına azalmış duyarlılık ya da dirençle ilişkili varyantlar tanımlanmıştır ve uygun rejimler ile kalıcı virolojik yanıt elde edilemeyen olguların çoğunda saptanmaktadır3,32. Proteaz inhibitörlerine karşı direnç, genotip 1a ve 1b izolatlarında da genel olarak farklılık göstermektedir. Proteaz geninde R155 (ve bazı durumlarda A156) aminoasit değişimleri, genel olarak çapraz direnç oluştururken, yeni bileşiklerin bazıları R155 varyantlarına etkili olabilmektedir32. Telaprevir ve bokeprevir; V36, T54, V55, A156 ve V170 varyasyonlarını seçerken, simeprevir ve diğer kovalan olmayan inhibitörlerde D168 değişiklikleri belirgindir. Simeprevir ayrıca, Q80 ve S122 varyantlarını da seçmektedir. Genotip 1a izolatlarında daha sık olarak izlenen Q80K varyantı dışında, direnç varyantlarının prevalansı, tedavi almayan olgularda düşüktür ve tedavi öncesinde 1a ile enfekte olgularda bu varyantın test edilmesi önerilmektedir32. Sofosbuvir de, NS5B geninde S282T ve/veya L159F/L320F değişiklikleri izlenen dirençli varyantları seçmektedir33.
Eldeki veriler, proteaz inhibitörü direnci ile ilgili HCV varyantlarının, vahşi tip izolatlara göre azalmış replikasyon kapasitesine sahip olduğunu ve tedavi almamış olgularda nadiren izlendiğini göstermektedir32. Tedavi sonunda baskın halde olan dirençli virus popülasyonlarında, zaman içerisinde vahşi tip virusa geri dönüşüm saptanmıştır. Tedavi öncesi düşük sıklıkta bulunan dirençli mutantların, tedavi ile baskın hale geçişi, çeşitli olgu serilerinde gösterilmiştir. Bununla birlikte, dirençli varyantların varlığı, her koşulda yanıtsızlık anlamına gelmemekte; genotip farklılığı ve diğer faktörler, bu varyantların baskın hale gelişi ve tedaviye klinik yanıtı etkilemektedir32.
Viral mutantlara ek olarak, konağa ait bazı genetik değişikliklerin de, HCV enfeksiyonlarında tedavi yanıtı ile ilişkili olduğu ortaya konmuştur. İnterlökin(IL)-28B geninde, bazı tek nükleotid polimorfizmleri taşıyan, genotip 1b ve 2a ile enfekte olgular, IFN+ribavirin kombinasyon tedavisine olumlu yanıt vermemekte; bazı polimorfizmler ise enfeksiyonun kendiliğinden temizlenmesi ile ilişkilendirilmektedir. IL-28B gen polimorfizmleri DEA tedavisini de etkilemektedir32,34.
HCV GENOMİK VARYASYONLARININ SAPTANMASI
Standart dizileme ya da Sanger yöntemi ile dizileme, genomik varyasyonların saptanması amacıyla yaygın olarak kullanılmış; HCV epidemileri, genotipler, mutantlar, filogenetik modellemeler gibi birçok çalışma için veri sağlayan temel yöntem olmuştur4. Bununla birlikte, örnekte sadece baskın olan dizinin okunabilmesi ve türümsü havuzunda %15-20'den daha az yoğunlukla temsil edilen varyantların saptanamaması, bu yaklaşımın en önemli kısıtlamasıdır. Ayrıca, tüm varyant genomların ayrıntılı olarak analizi, hedeflerin klonlanması ve yüksek sayıda klonun analizi ile mümkün olabilmektedir. Yeni jenerasyon dizileme tekniklerinde (next generation sequencing, NGS) izlenen gelişmelerle, bu yaklaşımlar HCV çeşitliliğinin incelenmesi için yaygın olarak kullanılmaya başlamıştır35. NGS ile elde edilen yüksek okuma sayısı; farklı dokular, konaklar ya da hastalık evrelerinde genetik çeşitliliğin ayrıntılı şekilde değerlendirilmesine imkan vermektedir24,26,27. Viral popülasyonda %1 ya da daha az sıklıkta bulunan kaçış mutantları ya da dirençli mutantların saptanması da mümkün olabilmektedir. NGS teknolojileri ve analiz yöntemleri hala gelişme sürecindedir ve dizileme hatalarının gerçek varyasyonlardan sağlıklı biçimde ayrılabilmesi için yaklaşımlar geliştirilmektedir35.
SONUÇ
Replikasyon sürecinin direkt sonucu olarak ortaya çıkan HCV genetik çeşitliliği, enfekte bireylerde türümsüler olarak izlenmektedir. Hastalık süresince türümsü içeriğinde belirgin değişiklikler olmakta; bu sırada enfeksiyonun kronikleşmesi ve ilerleyişinden sorumlu olan kaçış mutantları seçilime uğramaktadır. Günümüzde HCV genomunda, konağın immün yanıtı ve antiviral tedavi uygulamaları sonucu varyasyon gösteren bölgeler ve kaçıştan sorumlu birçok mutasyon tanımlanmıştır. Genetik veriler, virusun zoonotik kökenlerine işaret etmiş, 20. yüzyılda çeşitli nedenlerle izlenen dünyaya yayılım eğilimleri belirlenmiştir. Yeni geliştirilmekte olan DEA'lar ve tedavi rejimlerinin optimizasyonunda, virusun genetik çeşitliliğine ait veriler önemli rol oynayacaktır. NGS-temelli tekniklerin yaygınlaşmasıyla, HCV enfeksiyonlarının seyri ve önlenmesinde, varyant popülasyonların etkileri daha belirgin olarak ortaya konulacaktır.
KAYNAKLAR
- Gower E, Estes C, Blach S, Razavi-Shearer K, Razavi H. Global epidemiology and genotype distribution of the hepatitis C virus infection. J Hepatol 2014; 61(Suppl 1): S45-57.
- Simmonds P, Becher P, Collett MS, et al. Family Flaviviridae, pp: 1003-20. In: King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (eds), Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses. 2011, 1st ed. Elsevier Academic Press, San Diego.
- Di Lello FA, Culasso AC, Campos RH. Inter and intrapatient evolution of hepatitis C virus. Ann Hepatol 2015; 14(4): 442-9.
- Abdelrahman T, Hughes J, Main J, McLauchlan J, Thursz M, Thomson E. Next-generation sequencing sheds light on the natural history of hepatitis C infection in patients who fail treatment. Hepatology 2015; 61(1): 88-97.
- Duffy S, Shackelton LA, Holmes EC. Rates of evolutionary change in viruses: patterns and determinants. Nat Rev Genet 2008; 9(4): 267-76.
- Gray RR, Parker J, Lemey P, Salemi M, Katzourakis A, Pybus OG. The mode and tempo of hepatitis C virus evolution within and among hosts. BMC Evol Biol 2011; 11: 131.
- Honda M, Beard MR, Ping LH, Lemon SM. A phylogenetically conserved stem-loop structure at the 5' border of the internal ribosome entry site of hepatitis C virus is required for cap-independent viral translation. J Virol 1999; 73(2): 1165-74.
- Farci P. New insights into the HCV quasispecies and compartmentalization. Semin Liver Dis 2011; 31(4): 356-74.
- Salemi M, Vandamme A. Hepatitis C virus evolutionary patterns studied through analysis of full-genome sequences. J Mol Evol 2002; 54(1): 62-70.
- Sun HY, Ou NY, Wang SW, et al. Novel nucleotide and amino acid covariation between the 5'UTR and the NS2/NS3 proteins of hepatitis C virus: bioinformatic and functional analyses. PLoS One 2011; 6(9): e25530.
- Xu Z, Fan X, Xu Y, Di Bisceglie AM. Comparative analysis of nearly full length hepatitis C virus quasispecies from patients experiencing viral breakthrough during antiviral therapy: clustered mutations in three functional genes, E2, NS2, and NS5a. J Virol 2008; 82(19): 9417-24.
- Yi M, Ma Y, Yates J, Lemon SM. Compensatory mutations in E1, p7, NS2, and NS3 enhance yields of cell culture-infectious intergenotypic chimeric hepatitis C virus. J Virol 2007; 81(2): 629-38.
- Galli A, Bukh J. Comparative analysis of the molecular mechanisms of recombination in hepatitis C virus. Trends Microbiol 2014; 22(6): 354-64.
- Simmonds P. The origin of hepatitis C virus. Curr Top Microbiol Immunol 2013; 369: 1-15.
- Scheel TK, Simmonds P, Kapoor A. Surveying the global virome: identification and characterization of HCV-related animal hepaciviruses. Antiviral Res 2015; 115: 83-93.
- Smith DB, Bukh J, Kuiken C, et al. Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: updated criteria and genotype assignment web resource. Hepatology 2014; 59(1): 318-27.
- Walker CM. Adaptive immunity to the hepatitis C virus. Adv Virus Res 2010; 78: 43–86.
- Cashman SB, Marsden BD, Dustin LB. The humoral immune response to HCV: understanding is key to vaccine development. Front Immunol 2014; 5: 550.
- Skums P, Bunimovich L, Khudyakov Y. Antigenic cooperation among intrahost HCV variants organized into a complex network of cross-immunoreactivity. Proc Natl Acad Sci U S A 2015; 112(21): 6653-8.
- Lavillette D, Morice Y, Germanidis G, et al. Human serum facilitates hepatitis C virus infection, and neutralizing responses inversely correlate with viral replication kinetics at the acute phase of hepatitis C virus infection. J Virol 2005; 79(10): 6023-34.
- Prentoe J, Jensen TB, Meuleman P, et al. Hypervariable region 1 differentially impacts viability of hepatitis C virus strains of genotypes 1 to 6 and impairs virus neutralization. J Virol 2011; 85(5): 2224-34.
- Neumann-Haefelin C, Thimme R. Success and failure of virus-specific T cell responses in hepatitis C virus infection. Dig Dis 2011; 29(4): 416-22.
- Plauzolles A, Lucas M, Gaudieri S. Hepatitis C virus adaptation to T-cell immune pressure. ScientificWorldJournal 2013; 2013: 673240.
- Bull RA, Luciani F, McElroy K, et al. Sequential bottlenecks drive viral evolution in early acute hepatitis C virus infection. PLoS Pathog 2011; 7(9): e1002243.
- Roingeard P. Hepatitis C virus diversity and hepatic steatosis. J Viral Hepat 2013; 20(2): 77-84.
- Farci P, Quinti I, Farci S, et al. Evolution of hepatitis C viral quasispecies and hepatic injury in perinatally infected children followed prospectively. Proc Natl Acad Sci U S A 2006; 103(22): 8475-80.
- Miura M, Maekawa S, Takano S, et al. Deep-sequencing analysis of the association between the quasispecies nature of the hepatitis C virus core region and disease progression. J Virol 2013; 87(23): 12541-51.
- Li H, Sullivan DG, Feuerborn N, et al. Genetic diversity of hepatitis C virus predicts recurrent disease after liver transplantation. Virology 2010; 402(2): 248-55.
- Rupp D, Bartenschlager R. Targets for antiviral therapy of hepatitis C. Semin Liver Dis 2014; 34(1): 9-21.
- Khaliq S, Latief N, Jahan S. Role of different regions of the hepatitis C virus genome in the therapeutic response to interferon-based treatment. Arch Virol 2014; 159(1): 1-15.
- Bittar C, Jardim AC, Yamasaki LH, et al. On hepatitis C virus evolution: the interaction between virus and host towards treatment outcome. PLoS One 2013; 8(4): e62393.
- Kieffer TL, George S. Resistance to hepatitis C virus protease inhibitors. Curr Opin Virol 2014; 8: 16-21.
- Tong X, Le Pogam S, Li L, et al. In vivo emergence of a novel mutant L159F/L320F in the NS5B polymerase confers low-level resistance to the HCV polymerase inhibitors mericitabine and sofosbuvir. J Infect Dis 2014; 209(5): 668-75.
- Rauch A, Kutalik Z, Descombes P, et al. Genetic variation in IL28B is associated with chronic hepatitis C and treatment failure: a genome-wide association study. Gastroenterology 2010; 138(4): 1338-45.
- Quinones-Mateu ME, Avila S, Reyes-Teran G, Martinez MA. Deep sequencing: becoming a critical tool in clinical virology. J Clin Virol 2014; 61(1): 9-19.
İletişim (Correspondence):
Doç. Dr. Koray Ergünay,
Hacettepe Üniversitesi Tıp Fakültesi,
Tıbbi Mikrobiyoloji Anabilim Dalı,
06100 Sıhhiye, Ankara, Türkiye.
Tel (Phone): +90 312 305 1560,
E-posta (E-mail): ekoray@hacettepe.edu.tr